=== Hipótesis antropogénica o de caza excesiva ===
=== Hipótesis antropogénica o de caza excesiva ===
Por eso una de las hipótesis de creciente aceptación entre la comunidad científica es que estas extinciones masivas pudieron ser debidas fundamentalmente a la acción directa o indirecta del hombre<ref name=":0" />. Un reciente estudio estadístico que aplicaba modelos matemáticos para comparar la probabilidad de influencias de las dos variables (llegada humana y variación climática), ofreció como resultado que los fenómenos de extinción estaban más cerca de lo esperado por efecto del azar, al momento de la llegada humana en 76 casos (cada caso es la extinción de un género en una de las 19 regiones estudiadas). Por otro lado, el momento de la extinción fue más cercano al esperado por efecto del azar a los periodos de intenso cambio climático en sólo 2 casos (un valor cercano al esperado para una ordenación aleatoria de las extinciones a lo largo del tiempo). Las extinciones estaban más cerca de lo esperado por efecto del azar tanto a la llegada humana como a épocas de intensa variación climática en 8 casos; y a ninguna de estas variables en 40 casos, de un total de 126 casos. Estos resultados proporcionan un claro apoyo a la hipótesis de que los impactos antropogénicos, en lugar de la variación del clima, fueron el principal impulsor de las grandes extinciones cuaternarias de megafauna (76 casos frente a 2 de 126 casos analizados)<ref>{{Cita web|url=https://www.researchgate.net/publication/230623983_Human_arrival_scenarios_have_a_strong_influence_on_interpretations_of_the_late_Quaternary_extinctions?el=1_x_8&enrichId=rgreq-16602a991402369d07b9bdaa63d27f85-XXX&enrichSource=Y292ZXJQYWdlOzMwNTQ4OTE3NjtBUzozODYzMzYzNDAxMDMxNjhAMTQ2OTEyMTE2OTAyNA==|título=Human arrival scenarios have a strong influence on interpretations of the late Quaternary extinctions (PDF Download Available)|fechaacceso=2017-02-21|idioma=en|sitioweb=ResearchGate}}</ref>.
Por eso una de las hipótesis de creciente aceptación entre la comunidad científica es que estas extinciones masivas pudieron ser debidas fundamentalmente a la acción directa o indirecta del hombre<ref name="sudden death">{{cita libro|título=American Megafaunal Extinctions at the End of the Pleistocene|año=2009|isbn=978-1-4020-8792-9|doi=10.1007/978-1-4020-8793-6_2|páginas=21-37|capítulo=Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction|series=Vertebrate Paleobiology and Paleoanthropology|apellido1=Fiedel|nombre1=S.}}</ref><ref name=":2">{{cita publicación|título=Redefining the Age of Clovis: Implications for the Peopling of the Americas|nombre2=T. W.|publicación=Science|volumen=315|número=5815|páginas=1122-1126|bibcode=2007Sci...315.1122W|doi=10.1126/science.1137166|año=2007|apellido1=Waters|nombre1=M. R.|apellido2=Stafford}}</ref><ref name=":1">{{Cita web|url=https://www.researchgate.net/publication/305489176_Human_Dispersal_and_Late_Quaternary_Megafaunal_Extinctions_the_Role_of_the_Americas_in_the_Global_Puzzle|título=Human Dispersal and Late Quaternary Megafaunal Extinctions: the Role of the Americas in the Global Puzzle (PDF Download Available)|fechaacceso=2017-02-23|idioma=en|sitioweb=ResearchGate}}</ref><ref name=":4">{{Cita web|url=https://www.researchgate.net/publication/230623983_Human_arrival_scenarios_have_a_strong_influence_on_interpretations_of_the_late_Quaternary_extinctions?el=1_x_8&enrichId=rgreq-16602a991402369d07b9bdaa63d27f85-XXX&enrichSource=Y292ZXJQYWdlOzMwNTQ4OTE3NjtBUzozODYzMzYzNDAxMDMxNjhAMTQ2OTEyMTE2OTAyNA==|título=Human arrival scenarios have a strong influence on interpretations of the late Quaternary extinctions (PDF Download Available)|fechaacceso=2017-02-23|idioma=en|sitioweb=ResearchGate}}</ref><ref name=":02">{{cita publicación|url=http://www.uwyo.edu/surovell/pdfs/qi%202008.pdf|título=How many elephant kills are 14?: Clovis mammoth and mastodon kills in context|apellidos=|nombre2=N. M.|fecha=15 de noviembre de 2008|publicación=[[Quaternary International]]|editorial=|volumen=191|número=1|páginas=82–97|fechaacceso=|formato=|bibcode=2008QuInt.191...82S|doi=10.1016/j.quaint.2007.12.001|pmid=|apellido1=Surovell|nombre1=T. A.|apellido2=Waguespack|jstor=}}</ref><ref name=":5">{{Cita web|url=https://www.researchgate.net/publication/313113052_Paleoindians_Proboscideans_and_Phytotoxins_Exploring_the_Feasibility_of_Poison_Hunting_During_the_Last_Glacial-Interglacial_Transition|título=Paleoindians, Proboscideans, and Phytotoxins: Exploring the Feasibility of Poison Hunting During the Last Glacial-Interglacial Transition|fechaacceso=2017-02-23|idioma=en|sitioweb=ResearchGate}}</ref><ref name=":7">{{Cita web|url=https://www.researchgate.net/publication/305687531_Saving_the_World's_Terrestrial_Megafauna|título=Saving the World's Terrestrial Megafauna (PDF Download Available)|fechaacceso=2017-02-24|idioma=en|sitioweb=ResearchGate}}</ref><ref name=":8">{{Cita web|url=https://www.researchgate.net/publication/256669183_The_shifted_baseline_Prehistoric_defaunation_in_the_tropics_and_its_consequences_for_biodiversity_conservation|título=The shifted baseline: Prehistoric defaunation in the tropics and its consequences for biodiversity conservation (PDF Download Available)|fechaacceso=2017-02-24|idioma=en|sitioweb=ResearchGate}}</ref><ref name=":0" />. Un reciente estudio estadístico que aplicaba modelos matemáticos para comparar la probabilidad de influencias de las dos variables (llegada humana y variación climática), ofreció como resultado que los fenómenos de extinción estaban más cerca de lo esperado por efecto del azar, al momento de la llegada humana en 76 casos (cada caso es la extinción de un género en una de las 19 regiones estudiadas). Por otro lado, el momento de la extinción fue más cercano al esperado por efecto del azar a los periodos de intenso cambio climático en sólo 2 casos (un valor cercano al esperado para una ordenación aleatoria de las extinciones a lo largo del tiempo). Las extinciones estaban más cerca de lo esperado por efecto del azar tanto a la llegada humana como a épocas de intensa variación climática en 8 casos; y a ninguna de estas variables en 40 casos, de un total de 126 casos. Estos resultados proporcionan un claro apoyo a la hipótesis de que los impactos antropogénicos, en lugar de la variación del clima, fueron el principal impulsor de las grandes extinciones cuaternarias de megafauna (76 casos frente a 2 de 126 casos analizados)<ref>{{Cita web|url=https://www.researchgate.net/publication/230623983_Human_arrival_scenarios_have_a_strong_influence_on_interpretations_of_the_late_Quaternary_extinctions?el=1_x_8&enrichId=rgreq-16602a991402369d07b9bdaa63d27f85-XXX&enrichSource=Y292ZXJQYWdlOzMwNTQ4OTE3NjtBUzozODYzMzYzNDAxMDMxNjhAMTQ2OTEyMTE2OTAyNA==|título=Human arrival scenarios have a strong influence on interpretations of the late Quaternary extinctions (PDF Download Available)|fechaacceso=2017-02-21|idioma=en|sitioweb=ResearchGate}}</ref>.
==== Cronología de las extinciones masivas de megafauna y de la expansión de ''Homo sapiens'' ====
==== Cronología de las extinciones masivas de megafauna y de la expansión de ''Homo sapiens'' ====
En zoología terrestre, el término megafauna (del griego antiguomegas "grande" + el latínfauna "animal") se refiere a animales "gigantes", "muy grandes" o "grandes". El umbral más comúnmente usado es 44 kilogramos[1][2] o 100 kilogramos.[2][3] Por lo tanto esto incluye a muchas especies que no son popularmente consideradas como especialmente grandes, como el ciervo de cola blanca y el canguro rojo, e incluso a los humanos, aunque también se utiliza el criterio más restrictivo de aplicarlo a aquellos animales cuya masa excede una tonelada métrica (un millón de gramos o megagramo).[4]
En la práctica, un uso común encontrado en los escritos académicos y de divulgación se refiere a animales terrestres mayores que un ser humano y que no están (solamente) domesticados. El término es especialmente asociado con la megafauna del Pleistoceno y Holoceno que ha venido extinguiéndose en un proceso conocido como extinción masiva del Cuaternario desde hace 50.000 años hasta hoy— los animales grandes o gigantescos que se consideran arquetípicos de la última era de hielo como por ejemplo los mamutes en el hemisferio norte y los gliptodontes en América del Sur.[5] También se usa para referirse a los animales salvajes de mayor tamaño que existen, especialmente los elefantes, las jirafas, los hipopótamos, los rinocerontes, los alces, los cóndores, etc. La megafauna puede ser subcategorizada por su posición trófica en megaherbívoros (por ej. ciervos), megacarnívoros (leones), y más raramente, megaomnívoros (osos).
Otros usos comunes recaen en las especies gigantes acuáticas, como las ballenas, cualquier animal terrestre salvaje o domesticado como los antílopes y los toros, y los dinosaurios y otros reptiles gigantes extintos.
El término se aplica a animales (usualmente extintos) de gran tamaño parientes de un tipo de animal más común o sobreviviente, por ejemplo las libélulas de 1 metro de envergadura del período Carbonífero.
Estrategia ecológica
Megafauna —en el sentido de los mayores aves y mamíferos— son animales generalmente estrategas de la K, con una prolongada longevidad, una baja tasa de crecimiento poblacional, bajas tasas de mortandad, y pocos o ningún depredador capaces de matar a un adulto. Estas características, aunque no son exclusivas de dicha megafauna, los hacen altamente vulnerables a la sobreexplotación humana, lo que se cree que la habría podido llevar a fenómenos de extinción masiva en todos los continentes a excepción del Africa subsahariana y Asia meridional, los únicos lugares del planeta que conservan hoy en día una parte importante de la megafauna terrestre habitual en la mayor parte de los ecosistemas antes de nuestra expansión durante el Pleistoceno[6].
Evolución del gran tamaño corporal
Una observación que se ha hecho acerca de la evolución del tamaño corporal grande, dice que las tasas rápidas de incremento que se observan frecuentemente en intervalos relativamente cortos de tiempo no son sostenibles durante periodos mucho más prolongados. En un examen de las masas corporales de los mamíferos a través del tiempo, el máximo incremento posible en un intervalo de tiempo dado encontrado fue una escala con la longitud del intervalo elevado a la potencia 0,25.[7] Se cree que esto refleja la aparición, durante una tendencia de aumentar el tamaño máximo del cuerpo, de una serie de restricciones anatómicas, fisiológicas, ambientales, genéticas y de otro tipo que deben ser superadas por las innovaciones evolutivas antes de que posteriores incrementos de tamaño sean posibles. Una tasa de cambio sorprendentemente rápida se encontró para grandes disminuciones en la masa corporal, como la que pueden estar asociadas con el fenómeno del enanismo insular. Se encontró que cuando se normaliza la longitud de la generación, la tasa máxima de grandes reducciones de masa corporal es 30 veces mayor que la tasa máxima para los grandes incrementos de masa corporal.[7]
En mamíferos terrestres
Tras la extinción masiva del Cretácico-Terciario que eliminó a los dinosaurios hace cerca de 66 millones de años, los mamíferos terrestres experimentaron un casi exponencial aumento del tamaño corporal a medida que se diversificaban para ocupar los nichos ecológicos que quedaron vacantes.[8] Comenzando desde solo unos kilogramos antes de la extinción, su tamaño máximo se incrementó a 50 kg unos pocos millones de años después, y llega a 750 kg a finales del Paleoceno. Esta tendencia de aumento de la masa corporal parece estabilizarse hace cerca de 40 millones de años (en el Eoceno tardío), lo que sugiere que habían alcanzado restricciones ecológicas o fisiológicas, después de un incremento de masa corporal de tres órdenes de magnitud.[8] Sin embargo, si se considera desde el punto de vista de la tasa de aumento del tamaño de cada generación, se encuentra que el aumento exponencial continuó hasta la aparición de Paraceratherium hace 30 millones de años. (Dada las escalas de tiempo de generación de masa corporal0,259 , aumentando los tiempos de generación con el aumento de tamaño causan que el registro de la masa en función del tiempo forme una curva a la baja desde un ajuste lineal.)[7]
Los megaherbívoros eventualmente obtuvieron una masa corporal de más de 10.000 kg. Los mayores de estos, los indricoterios y los proboscídeos eran fermentadores del intestino grueso, que se cree que tenían una ventaja sobre los fermentadores del intestino delgado en términos de ser capaces de acelerar el tránsito gastrointestinal para poder acomodar la ingesta de enormes cantidades de comida.[9] Una tendencia similar aparece cuando se comparan las tasas de máximas masas corporales por generación para distintos clados de mamíferos (usando tasas promediadas sobre las escalas de tiempo macroevolutivas). Entre los mamíferos terrestres, las tasas más rápidas de incremento aparecen en los perisodáctilos, seguidos por los roedores y proboscídeos,[7] todos los cuales son fermentadores del intestino grueso. La tasa de incremento para los artiodáctilos fue cerca de un tercio de la de los perisodáctilos. La tasa para los carnívoros fue levemente menor, mientras que los primates, quizás restringidos por sus hábitos arborícolas, tuvieron la tasa más baja entre los grupos de mamíferos estudiados.[7]
Los mamíferos terrestres carnívoros de varios grupos de euterios (el mesoniquioAndrewsarchus, los creodontosMegistotherium y Sarkastodon, y los carnívorosAmphicyon y Arctodus) alcanzaron un tamaño máximo de cerca de 1.000 kg[8] (aparentemente Arctotherium era algo más grande). El mayor carnívoro metaterio, Proborhyaena gigantea, aparentemente alcanzaba 600 kg, también cerca de este límite.Error en la cita: La etiqueta de apertura <ref> es incorrecta o tiene el nombre mal Un límite de tamaño máximo teórico ha sido predicho basándose en la tasa metabólica de los mamíferos, el costo energético de obtener las presas, y la tasa máxima estimada de consumo de presas.[10] También se ha sugerido que el tamaño máximo de los mamíferos carnívoros se ve restringido por el esfuerzo que el húmero puede resistir durante la máxima velocidad a la carrera.[11]
El análisis de la variación del tamaño corporal máximo en los últimos 40 millones de años sugiere que la caída de las temperaturas y el incremento del área de tierras continentales están asociados con el incremento del tamaño. La primera correlación podría ser consistente con la regla de Bergmann,[12] y podría estar relacionada con la ventaja termorregulatoria de una mayor masa corporal en los climas fríos,[8] la mejor habilidad de los organismos grandes para hacer frente a la disponibilidad de comida,[12] u otros factores;[12] la última correlación podría ser explicable en términos de limitaciones de rango y recursos.[8] Sin embargo, los dos parámetros están interrelacionados (debido a las caídas en el nivel del mar que acompañan el incremento de las glaciaciones), lo que hace que la razón principal de las tendencias de aumento de tamaño sean más difíciles de identicar.[8]
En aves no voladoras
Durante el Paleoceno, debido al tamaño pequeño inicial de los mamíferos, los nichos de superpredador fueron frecuentemente ocupados por miembros de otros grupos, como los crocodilianos terrestres (como Pristichampsus), grandes serpientes (Titanoboa), lagartos varánidos, o aves no voladoras[8] (como Gastornis en Europa y Norteamérica, Paleopsilopterus en Suramérica). En los continentes del norte, las grandes aves depredadores fueron desplazadas cuando evolucionaron los grandes carnívoros euterios. En la aislada América del Sur, los forusrácidos no pudieron ser desbancadas por los carnívoros metaterios esparasodontes locales y siguieron siendo dominantes hasta que los depredadores euterios llegaron desde Norteamérica (como parte del Gran Intercambio Biótico Americano) durante el Plioceno. Sin embargo, ninguna de las mayores aves depredadoras (Brontornis), posiblemente omnívoras (Dromornis[13]) o herbívoras (Aepyornis) del Cenozoico crecieron más allá de los 500 kg, y por lo tanto nunca alcanzaron el tamaño de los mayores depredadores mamíferos, sin mencionar el de los mayores mamíferos herbívoros. Se ha sugerido que el incremento en el grosor de las cáscaras de los huevos de ave en proporción a masa del huevo con el incremento en el tamaño del mismo implica un límite máximo para el tamaño de las aves.[14] La mayor especie de Dromornis, D. stirtoni, puede haberse extinguido después de haber obtenido la masa corporal aviana máxima y fue después desplazada por los marsupiales diprotodontes que evolucionaron a tamaños varias veces mayores.[15]
Extinciones en masa del Cuaternario
La extinción en masa de megafauna conocida como la extinción del Holoceno hoy tiende a encuadrarse por numerosos científicos dentro del evento de extinción del Cuaternario. Estos eventos de extinción masiva de megafauna comenzaron hace más de 45.000 años, durante el Pleistoceno, en Oceanía, tuvieron otro máximo al final del último período glacial (la glaciación de Würm) cuando se extinguieron muchos de los animales gigantes de la región holártica, como el mamut lanudo, en América y el norte de Eurasia, y continuó después en islas y archipiélagos de todo el mundo, siendo la última gran extinción masiva de megafauna la de Nueva Zelanda hace entre 900 y 500 años[16]. Australia perdió el 86% de su megafauna, la mayor parte entre hace 50.000 y 40.000 años[17]. En el caso de ambas Américas la extinción masiva afectó a decenas de especies de megafauna, con una extinción que afectó al 73% de las especies de megafauna en Norteamérica[18]. Así, desde hace al menos 50,000 años y comenzando por Australia, aproximadamente 2/3 de todos los animales terrestres mayores de 50 kilogramos se han extinguido en Eurasia, norte de África, Oceanía y las Américas[19]. Varias teorías han intentado atribuir las diversas olas de extinciones de megafauna de los últimos 50.000 años a causas no antropogénicas como cambios climáticos, enfermedades desconocidas (ninguna enfermedad conocida ha causado la extinción simultánea de géneros enteros pertenecientes a familias, órdenes e incluso infraclases totalmente distintas discriminando especies en función de su tamaño), un posible impacto extraterrestre (no registrado por los geólogos), entre otras. [20] [21][22][23]
Hipótesis climáticas
Las hipótesis climáticas, que fueron dominantes durante la mayor parte del pasado SXX, no son capaces de explicar por sí solas estos patrones de extinción masiva por varias razones:
• Hubo docenas de otros ciclos glaciales a lo largo del Pleistoceno, sin extinciones masivas asociadas en el registro fósil. Las extinciones en Australia y las islas del planeta no coincidíeron con los picos glaciales.
• Las extinciones presentan una selectividad de tamaño sin precedentes en el registro fósil, ya que afectan casi exclusivamente a especies de megafauna, cuando son las especies más pequeñas las más vulnerables a las variaciones de temperatura[24].
• Los fósiles muestran en el periodo inmediatamente anterior a la extinción una reducción drastica en la edad de madurez sexual de las especies de megafauna, que no se corresponde con lo registrado en extinciones debida a factores ambientales, pero que sí se ha estudiado y comprobado en repetidas ocasiones en poblaciones de mamíferos sujetos a una elevada presión cinegética[25].
• Por último los cambios climáticos no pueden explicar por qué las extinciones han sido sistemáticamente más recientes en las islas que en los continentes, cuando ante un cambio climatico lo esperado es justo lo contrario: las poblaciones insulares, sin posibilidades de emigrar como las continentales, y con un número de ejemplares y un pool genético mucho menores, debieron extinguirse en primer lugar. Por el contrario, sí parece existir una correspondencia cada vez más clara entre las fechas de llegada de los humanos con las de la extinción de la megafauna en cada masa terrestre[19].
El enfoque global de las extinciones nos proporciona "experimentos naturales" en el tiempo y el espacio que nos permiten separar los factores climáticos de varias extinciones masivas: la megafauna sobrevivió a través de muchos otros ciclos glaciales intensos a lo largo de todo el Pleistoceno, como se ha comentado, pero además también persistió hasta bien entrado el Holoceno en islas que habían permanecido fuera del alcance de nuestra especie (hasta hace 4000 años en el caso de los mamuts siberianos en islas del Ártico o de los perezosos gigantes en las Antillas del Caribe, y hasta el SXVIII en el caso de la vaca marina de Steller en el Mar de Bering) y su extinción además parece coincidir con los primeros registros humanos en todas estas estas islas. Ambas líneas de evidencias muestran que si no fuera por los impactos antropogénicos, es poco probable que la megafauna se hubiera extinguido. Por lo tanto, en la mayor parte de los casos de extinción masiva de megafauna los cambios climáticos parecen haber actuado principalmente como intensificadores, cuando coincideron con la llegada de los humanos a una masa terrestre determinada.[26].
Hipótesis antropogénica o de caza excesiva
Por eso una de las hipótesis de creciente aceptación entre la comunidad científica es que estas extinciones masivas pudieron ser debidas fundamentalmente a la acción directa o indirecta del hombre[27][28][29][30][31][32][33][34][19]. Un reciente estudio estadístico que aplicaba modelos matemáticos para comparar la probabilidad de influencias de las dos variables (llegada humana y variación climática), ofreció como resultado que los fenómenos de extinción estaban más cerca de lo esperado por efecto del azar, al momento de la llegada humana en 76 casos (cada caso es la extinción de un género en una de las 19 regiones estudiadas). Por otro lado, el momento de la extinción fue más cercano al esperado por efecto del azar a los periodos de intenso cambio climático en sólo 2 casos (un valor cercano al esperado para una ordenación aleatoria de las extinciones a lo largo del tiempo). Las extinciones estaban más cerca de lo esperado por efecto del azar tanto a la llegada humana como a épocas de intensa variación climática en 8 casos; y a ninguna de estas variables en 40 casos, de un total de 126 casos. Estos resultados proporcionan un claro apoyo a la hipótesis de que los impactos antropogénicos, en lugar de la variación del clima, fueron el principal impulsor de las grandes extinciones cuaternarias de megafauna (76 casos frente a 2 de 126 casos analizados)[35].
Cronología de las extinciones masivas de megafauna y de la expansión de Homo sapiens
El último gran pulso de extinción cerca del final del Pleistoceno e inicio del Holoceno fue por tanto sólo uno más de una serie de extinciones masivas de megafauna que han ocurrido desde hace 50.000 años hasta el tiempo actual en el que la megafauna superviviente de África y Asia, está viendo también sus poblaciones drásticamente reducidas. Dentro del actual ciclo de extinción masiva en que nos encontramos, en el periodo que va desde 1970 a 2012, y de acuerdo con un análisis global reciente, ha habido una disminución general del 58% en los efectivos de las poblaciones de vertebrados[36]. Todas las extinciones masivas anteriores parecen haber coincidido en el tiempo con la expansión del ser humano moderno a nuevos continentes o grandes islas, siendo el África subsahariana y el sur de Asia las regiones menos afectadas por el proceso hasta el SXIX[37]. Se apunta a que esto puede ser debido a que ambas zonas soportaron una larga presencia de nuestro género desde su aparición. Así estas especies de megafauna pudieron coevolucionar con especies del género Homo y evitar instintivamente su contacto o desarrollar respuestas agresivas a su presencia, y lo habrían hecho ya bastante antes de que surgiera nuestra especie, más efectiva aún que las anteriores especies de Homo como cazador. La megafauna del resto del mundo no habría desarrollado mecanismos instintivos para identificarnos como depredador, un comportamiento que se conoce como ingenuidad insular o ingenuidad ecológica (island tameless o island naivety en inglés) y que se ha registrado en numerosas ocasiones (Diamond, 1984) y aún hoy se observa en aquellas especies endémicas de islas y continentes de reciente descubrimiento como las Galápagos o la Antártida [38] [39]. Este comportamiento es identificado por los ecólogos como un factor clave de vulnerabilidad de las especies insulares frente a la aparición de especies alóctonas[40]. Así por ejemplo los pingüinos del continente antártico muestran un fuerte instinto de huida frente a especies depredadoras con las que han coevolucionado, como la foca leopardo, pero no frente al ser humano, al que no temen.
Fuera del territorio continental de Afro-Eurasia, estas extinciones de megafauna siguieron un patrón distintivo de masa terrestre por masa terrestre que seguía a la expansión humana en regiones previamente no habitadas del globo, lo que como se ha expuesto no muestra correlación con la historia climática (la cual puede ser visualizada con gráficos sobre periodos de tiempos geológicos recientes de marcadores climáticos como isótopos de oxígeno marino o niveles de dióxido de carbono atmosférico).[41][42] Australia fue afectada primero hace cerca de 45.000 años,[43] seguida por Tasmania hace cerca de 41.000 años (después de la formación de un puerte terrestre con Australia hace unos 43.000 años),[44][45][46] Japón aparentemente hace cerca de 30.000 años,[47] Norteamérica hace 13.000 años, Sudamérica cerca de 500 años después,[48][49] Chipre hace 10.000 años,[50][51] las Antillas hace 6.000 años,[52] Nueva Caledonia[53] y las islas cercanas[54] hace 3.000 años, Madagascar hace 2.000 años,[55] Nueva Zelanda hace 700 años,[56] las islas Mascarenas hace 400 años,[57] y las islas del Comandante hace 250 años.[58] Casi todas las islas aisladas del mundo podrían proporcionar ejemplos similares de extinciones que ocurrieron poco después de la llegada de Homo sapiens, aunque muchas de estas islas, como Hawaii, nunca tuvieron una megafauna terrestre, por lo que su fauna extinta era menor.[41][42]
Un análisis de las esporas del hongo Sporormiella (que se derivan mayormente del excremento de los megaherbívoros) en núcleos de sedimentos de pantano que abarcan los últimos 130.000 años del cráter de Lynch en Queensland, Australia muestran que la megafauna había desaparecido virtualmente hace cerca de 41.000 años, en una época en la que los cambios climáticos eran mínimos; el cambio fue acompañado por el incremento del carbón de origen vegetal, y fue seguido por la transición de las selvas a la vegetación esclerófila resistente al fuego. La alta resolución de la cronología de los cambios apoya la hipótesis de que sólo la caza por parte de los humanos eliminó la megafauna, y que el subsiguiente cambio en la flora fue muy probablemente una consecuencia de la eliminación de los ramoneadores y del incremento de los incendios[59][60][61] El incremento de los incendios siguió a la desaparición de la megafauna por cerca de un siglo, y muy probablemente resultó de la acumulación de material combustible cuando se detuvo el pastoreo. En los siglos siguientes los pastos se incrementaron; la vegetación esclerófila se incrementó con un intervalo de otro siglo, y el bosque de esclerófilas se desarrolló después de otros mil años.[61] Durante dos períodos de cambio climático entre hace 120 a 75 mil años, la vegetación esclerófila también se había incrementado en el sitio en respuesta a las condiciones más frías y secas; ninguno de estos episodios tuvo un impacto significativo en la abundancia de megafauna.[61]
Extinciones masivas anteriores a la expansión de Homo sapiens
Varias extinciones en masa habían ocurrido antes en la historia geológica de la Tierra, en la cual se produjeron pérdidas de megafauna. La más conocida es la extinción del límite Cretácico-Terciario en la que la mayoría de los dinosaurios y varios reptiles gigantes fueron eliminados. Sin embargo, las extinciones en masa anteriores fueron más globales y no fueron tan selectivas para la megafauna; es decir, muchas especies de otros grupos, incluyendo plantas, invertebrados marinos[62] y plancton, se extinguieron también. Por tanto, los eventos anteriores deben haber sido causados por trastornos más generalizados en la biosfera que la irrupción de un nuevo factor local en cada una de las extinciones por continentes de los últimos 50.000 años.
Consecuencias de la extinción masiva de la megafauna
La extinción de la megafauna en buena parte de Eurasia, en las Américas y en Oceanía tuvo efectos devastadores y catastróficos para los ecosistemas terrestres, que llegan hasta nuestros días y cuyas dimensiones en perdidas de biodiversidad y de ecosistemas solo se están empezando a dilucidar[63].
Efectos sobre la biodiversidad y capacidad de carga de los ecosistemas
Con la extinción de la megafauna, la relación entre las diferentes especies cambia, hasta el punto de que varias especies de plantas se extinguen, y los animales supervivientes se ven afectados de muchas maneras. Tras la extinción de la megafauna se ha comprobado un aumento en la acumulación de la masa vegetal que eventualmente acaba provocando grandes incendios y cambios severos en la relación entre las especies, su diversidad y la capacidad de carga del ecosistema. Tal vez la consecuencia más visible a día de hoy es la relativa ausencia de animales grandes en relación a ecosistemas que conservan la megafauna original, incluso en áreas que son abandonadas durante miles de años por el hombre tras la extinción de la megafauna.
A modo de ejemplo se puede comparar las cifras de parques nacionales de Norteamérica deshabitados en la actualidad y África. En Canadá el Parque Nacional Jasper contiene aproximadamente 1300 wapitis y 150 alces. El parque tiene aproximadamente 11,000 km cuadrados y está muy bien protegido de la caza furtiva. Por el contrario, el parque nacional Chobe de tamaño similar, en Botswana, contiene más de 50.000 elefantes.
El Serengeti es uno de los espacios naturales menos alterados del mundo a pesar del furtivismo que sufre. Es sólo 3 veces mayor que Jasper, pero contiene 1,7 millones de ñus, 250.000 cebras y 500.000 gacelas. Además de elefantes, hipopótamos, jirafas, etc. En términos de depredadores Jasper contiene aproximadamente 110 osos grizzly, 90 osos negros y sólo 45 lobos. El Serengeti tiene más de 3000 leones, 8000 hienas y 1000 leopardos. Estos números son aún más reseñables cuando se considera la considerable presión de la caza furtiva sobre la fauna africana. La población de búfalos cafre en el Serengeti era de 65.000 en 1969, pero se ha reducido a 16.000 en la actualidad.
Jasper tiene una temporada de crecimiento más corta, con menos intensa luz solar, mientras que el Serengeti sufre caza furtiva y una sequía estacional. Sin embargo, existe una enorme diferencia en la cantidad de animales que viven en ambos parques. Una razón fundamental de esta diferencia en el tamaño de las poblaciones es la reciente extinción de la megafauna en América del Norte.
Hoy es muy difícil estimar la cantidad de animales que estarían presentes si Jasper fuera aún un espacio intacto. Un estudio[64] de restos fósiles en Rusia proporciona algunas cifras para las praderas subárticas del norte de Siberia. Las conclusiones de la investigación sugieren aproximadamente 1 mamut por kilómetro cuadrado en uno de los ambientes más fríos de la tierra. También proporciona cifras para algunas otras especies de megafauna. Si se extrapolan las cifras, Jasper pudo albergar unos 11.000 mamuts, 66.000 caballos y 165.000 ciervos. En Jasper, el papel de los mamuts europeos se vería probablemente dividido entre los mamuts americanos y otro enorme elefante conocido como mastodonte. El área también debería contener camellos, perezosos gigantes, castores gigantes, leones americanos, jaguares, tigres de dientes de sable, antílopes saiga, lobos gigantes, osos gigantes, guepardos americanos, bisontes gigantes, varias especies de buey almizclero y berrendos.
Efectos en el transporte de nutrientes
La megafauna juega un papel importante en el transporte lateral de nutrientes minerales en los ecosistemas, tendiendo a traslocarlos de las áreas de gran abundancia a aquellas de menor cantidad. Este movimiento se debe al tiempo entre el consumo de los nutrientes y el tiempo en que los liberan a través de la excreción (o, en menor medida, a través de la descomposición tras su muerte).[65] Se estima que en la Cuenca amazónica de Suramérica, esta difusión lateral se redujo en un 98% tras las extinciones de la megafauna que ocurrieron hace aproximadamente 12,500 años.[66][67] Dado que se cree que la disponibilidad del fósforo limita la productividad en la mayor parte de la región, la disminución de este transporte desde la parte occidental de la cuenca y desde las planicies aluviales (ambas regiones que derivan su suministro de las elevaciones de los Andes a otras áreas debió de impactar significativamente la ecología de la región, y los efectos pueden no haber terminado de alcanzar su límite.[67]
Efectos en la dispersion de semillas
Algunas plantas que dependen de la megafauna para su dispersión de semillas, todavía están vivas y sin ella se están extinguiendo lentamente. Un ejemplo llamativo es el naranjo de Luisiana. Este árbol produce una fruta de gran tamaño que ningún animal americano come en la actualidad. Para la planta esto supone una enorme pérdida de energía, ya que el próposito de cualquier fruto es dispersar semillas a través de los animales que lo consumen. Como ningún animal moderno lo come, el fruto cae al suelo y las semillas quedan bajo el árbol. De este modo el rango de distribucion de estas plantas se contrae, en vez de extenderse. Cuando los europeos registraron por primera vez el naranjo de Luisiana, su área se limitaba a un pequeño territorio en el centro / norte de Texas. Sin embargo, en el registro polínico fósil aparece extendido por todo el continente. De hecho, hoy sabemos que había 7 especies, los otros 6 disminuyeron sus poblaciones hasta la extinción sin la megafauna. Estos árboles de frutas grandes deberían ser mucho más comunes de lo que son hoy debido a la extinción de la megafauna.
En Sudamérica un estudio estadístico del 2015 ha puesto de manifiesto que decenas de especies de árboles (más de 60) cuyos frutos se cree que eran dispersados por la megafauna, han visto drásticamente reducida su distribución desde la extinción de estos animales[68].
Efectos de la desaparición de la megafauna sobre las emisiones de metano
Muchos herbívoros producen metano como un subproducto de la fermentación en la digestión, y lo liberan eructando. Las grandes poblaciones de megafauna herbívora tiene el potencial de contribuir en gran medida a la concentración atmosférica de metano, el cual es un importante gas de invernadero. Hoy en día, cerca del 20% de las emisiones anuales de metano vienen del que libera el ganado. Estudios recientes han indicado que la extinción de los herbívoros de la megafauna puede haber causado una reducción del metano atmosférico. Esta hipótesis es relativamente nueva.[69]
Varios estudios han examinado el efecto de la eliminación de los mamíferos megaherbívoros en las emisiones de metano. Uno de estos examinó las emisiones de metano del bisonte que ocupaba las Grandes Planicies de Norteamérica antes de la llegada de los colonos europeos. Este estudio estimó que la desaparición de los bisontes causó un decrecimiento de 2.2 Tg/año. Este es un cambio proporcionalmente muy grande para este período de tiempo.[70]
Otro estudio examinó el cambio de la concentración de metano en la atmósfera al final del Pleistoceno después de la extinción de la megafauna del continente americano. Después de que los primeros humanos migraron a Américas hace más o menos 13.000 años antes del presente, su cacería y otros impactos ecológicos asociados llevaron a la extinción de muchas especies de megafauna en la región. Los cálculos sugieren que esta extinción produjo una caída en la producción de metano por ~9.6 Tg/año. Registros de núcleos de hielo apoyan esta hipótesis del rápido descenso de metano en esta época. Esto sugiere que la ausencia de emisiones de metano de la megafauna puede haber contribuido al abrupto enfriamiento climático al comienzo del Dryas Reciente.[69]
Ejemplos
Los siguientes son algunos ejemplos notables de animales frecuentemente considerados como megafauna (en el sentido de "animal grande"). Esta lista no pretende ser exhaustiva:
El canguro rojo (Macropus rufus) es el mayor mamífero viviente de Australia así como el mayor marsupial con un peso de 85 kg. Sin embargo, su pariente extinto, el canguro de cara corta Procoptodon alcanzaba 230 kg de peso, mientras que el extinto Diprotodon alcanzó el mayor tamaño de cualquier marsupial, estimado en más de 2.750 kg. El extinto león marsupial (Thylacoleo carnifex), con más de 160 kg era mucho mayor que cualquier marsupial carnívoro. Se extinguieron después de la llegada del hombre.
Los elefantes son los mayores animales terrestres vivientes. Estos y sus parientes cercanos se originaron en África, pero hasta hace poco tenían una distribución casi mundial. El elefante africano de sabana (Loxodonta africana) tiene una altura a los hombros de 4.3 metros y pesar más de 8 toneladas. Entre los proboscídeos recientemente extintos los mamuts (Mammuthus) eran parientes cercanos de los elefantes, mientras que los mastodontes (Mammut) estaban mucho menos relacionados. Se extinguieron poco depués de la llegada del hombre a América. Se estima que el mamut del río Songhua (M. sungari) pesaba hasta 17 toneladas, lo que lo convierte en uno de los mayores proboscídeos y el segundo mayor animal terrestre después de los indricoterinos.
El mayor sirenio moderno con más de 1.500 kg de peso es el manatí (Trichechus manatus). La vaca marina de Steller (Hydrodamalis gigas) era probablemente cinco veces más grande, pero infortunadamente fue exterminada por los humanos en los 27 años siguientes a su redescubrimiento en las remotas islas del Comandante en 1741. En época prehistórica este animal se reproducía también en las costas del noreste de Asia y el noroeste de Norteamérica; fue probablemente eliminado de estos lugares más accesibles por cazadores[71].
Los perezosos terrestres fueron otro grupo de lentos xenartros terrestres, emparentados con los actuales perezosos. Estos tuvieron una historia similar, aunque alcanzaron Norteamérica mucho antes, y se expandieron más al norte con Megalonyx. Los mayores géneros, Megatherium y Eremotherium, alcanzaban tamaños comparables al de los elefantes. Se extinguieron después de la llegada del hombre a América.
El actual capibara (Hydrochoerus hydrochaeris) de Suramérica, es el mayor roedor vivo, pesando más de 65 kg. Varias formas norteamericanas recientemente extintas eran mayores: el capibara Neochoerus pinckneyi (otro migrante neotropical) que era 40% más pesado; el castor gigante (Castoroides ohioensis) era similar. La extinta jutia gigante (Amblyrhiza inundata) de varias islas del Caribe puede haber sido incluso mayor. Sin embargo, hace varios millones de años Sudamérica alojaba roedores mucho más grandes. Phoberomys pattersoni, conocido de un esqueleto casi completo, probablemente alcanzaba 700 kg de peso. Restos fragmentarios sugieren que Josephoartigasia monesi crecía hasta más de 1.000 kg.
Los grandes felinos incluyen al tigre (Panthera tigris) y al león (Panthera leo). La mayor subespecie, con más de 306 kg, es el tigre siberiano (P. tigris altaica), de acuerdo con la regla de Bergmann. Los miembros del género Panthera se distinguen por características morfológicas que les permiten rugir. Entre los mayores felinos extintos se encuentra el león americano (Panthera leo atrox) y el dientes de sable suramericano Smilodon populator. Se extinguieron poco después de la llegada del hombre a América.
Los osos son los mayores carnívoros del suborden Caniformia. Las mayores formas vivientes son el oso polar (Ursus maritimus), con un peso de más de 680 kg, y el oso de Kodiak (Ursus arctos middendorffi) de tamaño similar, ambos también consistentes con la regla de Bergmann. Arctotherium angustidens, un oso de cara corta de Sudamérica, fue uno de los mayores mamíferos depredadores terrestres conocidos con un peso promedio estimado en 1.600 kg.[78]
Los pinípedos (morsas, focas y leones marinos) son carnívoros marinos anfibios que evolucionaron de ancestros similares a los osos. El elefante marino del sur (Mirounga leonina) de las aguas de la Antártida y subantárticas es el mayor de todos los carnívoros, con machos adultos alcanzando una longitud máxima de 6 a 7 metros y un peso máximo de 5.000 kilogramos.
Los tapires son animales ramoneadores, con una nariz prensil, algo similares a un cerdo que parece que han cambiado poco en los últimos 20 millones de años. Estos habitan las selvas tropicales del Sureste Asiático y de Centro y Suramérica, y están entre los mayores mamíferos terrestres existentes de las dos últimas regiones.
RinoceronteLos rinocerontes son perisodáctilos con cuernos hechos de queratina, el mismo tipo de proteína de la se compone el pelo. Estos están entre los mayores mamíferos terrestres vivientes después de los elefantes (los hipopótamos alcanzan un tamaño similar). Tres de la cinco especies actuales están en peligro crítico. Sus extintos parientes del Asia central los indricoterinos fueron los mayores mamíferos terrestres de todos los tiempos.
Los hipopótamos (Hippopotamus amphibius) son los artiodáctilos vivos más pesados; se cre que este y el amenazado] hipopótamo enano (Choeropsis liberiensis) son parientes cercanos de los cetáceos. Los hipopótamos africanos están estre las especies de megafauna más peligrosos para los humanos.[79]
Orden Cetacea (o cladísticamente, Cetartiodactyla)
Los Cetáceos (ballenas, delfines y marsopas) son mamíferos marinos. La ballena azul (Balaenoptera musculus) es el mayor de los rorcuales y el mayor animal que haya vivido. El cachalote (Physeter macrocephalus) es el mayor odontoceto, así como el animal más ruidoso y con mayor cerebro (que es cinco veces más grande que el de los seres humanos). La orca (Orcinus orca) es el delfín de mayor tamaño.
Los extintos dromornítidos de Australia como Dromornis pueden haber excedido a las mayores ratites en tamaño. Debido a su pequeño tamaño para un continente y su aislamiento, Australia es vista en ocasiones com la mayor isla del mundo; por tanto, estas especies podrías ser también consideradas como gigantes insulares.
Los dinosaurios saurisquios del Jurásico y el Cretácico incluyen a los saurópodos, los animales terrestres más largos (con más de 40 metros) y más pesados que se conozcan (Argentinosaurus alcanzaba entre 80–100 toneladas métricas, así como a los terópodos, los mayores carnívoros terrestres (Spinosaurus alcanzaba entre 7 a 9 toneladas).
Aunque el mayor lagarto viviente, el dragón de Komodo (Varanus komodoensis), otro gigante isleño, puede alcanzar 3 metros de largo, su pariente extinto australiano Megalania podía alcanzar más del doble de ese tamaño. Los parientes marinos extintos de los varánidos, los mosasáuridos, fueron superdepredadores en los mares del Cretácico Superior.
Se considera que la serpiente más pesada es la anaconda verde (Eunectes murinus), mientras que la pitón reticulada (Python reticulatus), con más de 8.7 metros, es la más larga. Una especie extinta australiana del Plioceno de Liasis, la L. dubudingala, alcanzaba 10 metros, mientras que Titanoboa del Paleoceno de Suramérica alcanzaba longitudes de 12–15 metros y un peso estimado de cerca de 1.135 kilogramos.
La mayor tortuga es la amenazada tortuga laúd (Dermochelys coriacea), que pesa más de 900 kg. Se distingue de otras tortugas marinas por su carencia de caparazón óseo. Los quelonios terrestres más grandes son las tortugas gigantes de las islas Galápagos (Chelonoidis nigra) y el atolón de Aldabra (Aldabrachelys gigantea), con más de 300 kg. Estas tortugas son los mayores sobrevivientes de una variedad de especies de tortugas gigantes que estuvieron ampliamente representadas en las masas terrestres continentales[80][81] y varias islas[80] durante el Pleistoceno.
El temnospóndilo del PérmicoPrionosuchus, el mayor anfibio conocido, alcanzaba 9 metros de largo y era un depredador acuático similar a un cocodrilo. Tras la aparición de los crocodilianos verdaderos, temnospóndilos como Koolasuchus (de 5 metros de largo) se retiraron a la región de la Antártida en el Cretácico, antes de extinguirse.
El mayor pez óseo es el pez luna (Mola mola), cuyo peso adulto en promedio es de 1.000 kg. Aunque filogenéticamente es un "pez óseo", su esqueleto es sobre todo cartílago (que es más ligero que el hueso). Tiene un cuerpo en forma de disco, y se impulsa con sus largas y delgadas aletas dorsales y anales; se alimenta principalmente de medusas. En estos tres aspectos (así como su tamaño y hábitos de buceo), se parece a la tortuga laúd.
El amenazado esturión europeo (Huso huso) con más de 1476 kg) es el esturión de mayor tamaño (los cuales son también mayormente cartilaginosos) y se considera el mayor pez anadromo.
El mayor pez depredador vivo, el gran tiburón blanco (Carcharodon carcharias), alcanza hasta 2.240 kg de peso. Su pariente extinto C. megalodon (se debate si pertenece al género Carcharodon o Carcharocles) era mayor por más de un orden de magnitud, y es el mayor tiburón y pez depredador de cualquier época; cazaba ballenas y otros mamíferos marinos.
El mayor tiburón actual, pez cartilaginoso y pez en general es el tiburón ballena (Rhincodon typus), que llega a pesar más de 21.5 toneladas. Como las ballenas, se alimenta por filtración y consume mayormente plancton.
Un cierto número de criaturas de las profundidades oceánicas exhiben gigantismo abisal. Estas incluyen al calamar gigante (Architeuthis) y al calamar colosal (Mesonychoteuthis hamiltoni); se cree que ambos cefalópodos (raramente vistos) alcanzan longitudes de 12 metros o más. El último es el invertebrado de mayor tamaño conocido, y tiene los mayores ojos de cualquier animal. Ambos son presas de los cachalotes.
Galería
Extintos
Fósil de un Arthropleura, una escolopendra gigante que vivió en el período Carbonífero.
↑Fariña, Richard A.; Vizcaíno, Sergio F.; De Iuliis, Gerry (2013). Megafauna. Giant Beasts of Pleistocene South America. Indiana University Press. p. 416. ISBN978-0-253-00230-3.
↑ abcdefgSmith, F. A.; Boyer, A. G.; Brown, J. H.; Costa, D. P.; Dayan, T.; Ernest, S. K. M.; Evans, A. R.; Fortelius, M.; Gittleman, J. L.; Hamilton, M. J.; Harding, L. E.; Lintulaakso, K.; Lyons, S. K.; McCain, C.; Okie, J. G.; Saarinen, J. J.; Sibly, R. M.; Stephens, P. R.; Theodor, J.; Uhen, M. D. (26 de noviembre de 2010). «The Evolution of Maximum Body Size of Terrestrial Mammals». Science330 (6008): 1216-1219. doi:10.1126/science.1194830. Consultado el 7 de enero de 2012.La referencia utiliza el parámetro obsoleto |coautores= (ayuda)
↑Fiedel, S. (2009). «Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction». American Megafaunal Extinctions at the End of the Pleistocene. Vertebrate Paleobiology and Paleoanthropology. pp. 21-37. ISBN978-1-4020-8792-9. doi:10.1007/978-1-4020-8793-6_2.
↑JOSÉ MARÍA FERNÁNDEZ-PALACIOS (2004). «INTRODUCCIÓN A LAS ISLAS». http://jmferpal.webs.ull.es. ECOLOGÍA INSULAR / ISLAND ECOLOGY. ASOCIACIÓN ESPAÑOLA DE ECOLOGÍA TERRESTRE (AEET). Consultado el 21 de febrero de 2017.
↑Anderson, A.; Sand, C.; Petchey, F.; Worthy, T. H. (2010). «Faunal extinction and human habitation in New Caledonia: Initial results and implications of new research at the Pindai Caves». Journal of Pacific Archaeology1 (1): 89-109. hdl: 10289/5404.La referencia utiliza el parámetro obsoleto |coautores= (ayuda)
↑White, A. W.; Worthy, T. H.; Hawkins, S.; Bedford, S.; Spriggs, M. (16 de agosto de 2010). «Megafaunal meiolaniid horned turtles survived until early human settlement in Vanuatu, Southwest Pacific». Proc. Natl. Acad. Sci. USA107 (35): 15512-15516. doi:10.1073/pnas.1005780107.La referencia utiliza el parámetro obsoleto |coautores= (ayuda)
↑O’Connor, M.; Wolf, A.; Doughty, C. E.; Malhi, Y. (9 de agosto de 2013). «Lateral Diffusion of Nutrients by Mammalian Herbivores in Terrestrial Ecosystems». PLoS ONE8 (8): e71352. doi:10.1371/journal.pone.0071352.
↑ abDoughty, C. E.; Wolf, A.; Malhi, Y. (11 de agosto de 2013). «The legacy of the Pleistocene megafauna extinctions on nutrient availability in Amazonia». Nature Geoscience. doi:10.1038/ngeo1895.




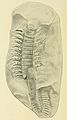 Fósil de un Arthropleura, una escolopendra gigante que vivió en el período Carbonífero.
Fósil de un Arthropleura, una escolopendra gigante que vivió en el período Carbonífero. Dunkleosteus era un pez placodermo acorazado depredador del Devónico.
Dunkleosteus era un pez placodermo acorazado depredador del Devónico.

 Tyrannosaurus era un terópodo que medía más de 12 m de largo, y era el superdepredador del oeste de Norteamérica.
Tyrannosaurus era un terópodo que medía más de 12 m de largo, y era el superdepredador del oeste de Norteamérica. Saurópodos macronarianos; de izquierda a derecha, Camarasaurus, Brachiosaurus, Giraffatitan y Euhelopus.
Saurópodos macronarianos; de izquierda a derecha, Camarasaurus, Brachiosaurus, Giraffatitan y Euhelopus.
 El C. megalodon es el mayor tiburón que se conoce.
El C. megalodon es el mayor tiburón que se conoce. Deinotherium tenía colmillos vueltos hacia bajo y estaba ampliamente distribuido en Afro-Eurasia.
Deinotherium tenía colmillos vueltos hacia bajo y estaba ampliamente distribuido en Afro-Eurasia. Titanis walleri, la única ave del terror que se sabe invadió América del Norte, medía 2 metros de alto.
Titanis walleri, la única ave del terror que se sabe invadió América del Norte, medía 2 metros de alto.

 Mylodon, un tipo de perezoso gigante (escultura ubicada en el Parque Pleistocénico de Osorno.)
Mylodon, un tipo de perezoso gigante (escultura ubicada en el Parque Pleistocénico de Osorno.) Megatherium, del Pleistoceno suramericano, era del tamaño de un elefante y fue el mayor perezoso terrestre.
Megatherium, del Pleistoceno suramericano, era del tamaño de un elefante y fue el mayor perezoso terrestre. Toxodon, uno de los mayores y últimos de los notoungulados. Parientes suyos se expandieron hasta Estados Unidos.
Toxodon, uno de los mayores y últimos de los notoungulados. Parientes suyos se expandieron hasta Estados Unidos. El león americano excedía el tamaño del actual león y vivieron en Norteamérica hasta hace 10.000 años antes del presente.
El león americano excedía el tamaño del actual león y vivieron en Norteamérica hasta hace 10.000 años antes del presente.![Los mamutes lanudos desaparecieron después de que los humanos invadieran su hábitat en Eurasia y América del Norte.[1]](//upload.wikimedia.org/wikipedia/commons/thumb/a/a3/Woolly_mammoth_%28Mammuthus_primigenius%29_-_Mauricio_Ant%C3%B3n.jpg/120px-Woolly_mammoth_%28Mammuthus_primigenius%29_-_Mauricio_Ant%C3%B3n.jpg) Los mamutes lanudos desaparecieron después de que los humanos invadieran su hábitat en Eurasia y América del Norte.[1]
Los mamutes lanudos desaparecieron después de que los humanos invadieran su hábitat en Eurasia y América del Norte.[1] El águila de Haast, la mayor águila conocida, atacando a una moa (que incluye a las más altas aves conocidas).
El águila de Haast, la mayor águila conocida, atacando a una moa (que incluye a las más altas aves conocidas).








.jpg)

.jpg)



![Los mamutes lanudos desaparecieron después de que los humanos invadieran su hábitat en Eurasia y América del Norte.[1]](/wiki/Archivo:Woolly_mammoth_(Mammuthus_primigenius)_-_Mauricio_Ant%C3%B3n.jpg)

 El gorila es el mayor primate y uno de los más seriamente amenazados en el planeta.
El gorila es el mayor primate y uno de los más seriamente amenazados en el planeta.
 El oso polar es el mayor oso y un carnívoro semiacuático, y son vulnerables al calentamiento global.
El oso polar es el mayor oso y un carnívoro semiacuático, y son vulnerables al calentamiento global. El rinoceronte negro, de más de 4 metros de largo, se encuentra amenazado por la caza furtiva.
El rinoceronte negro, de más de 4 metros de largo, se encuentra amenazado por la caza furtiva. Los camellos bactrianos salvajes están gravemente amenazados. Sus ancestros se originaron en Norteamérica.
Los camellos bactrianos salvajes están gravemente amenazados. Sus ancestros se originaron en Norteamérica.![A diferencia del rinoceronte y el mamut lanudos, el buey almizclero logró sobrevivir a las extinciones del Cuaternario.[1]](//upload.wikimedia.org/wikipedia/commons/thumb/e/ed/Ovibos_moschatus_qtl3.jpg/120px-Ovibos_moschatus_qtl3.jpg) A diferencia del rinoceronte y el mamut lanudos, el buey almizclero logró sobrevivir a las extinciones del Cuaternario.[1]
A diferencia del rinoceronte y el mamut lanudos, el buey almizclero logró sobrevivir a las extinciones del Cuaternario.[1] Un hipopótamo, El más pesado y más acuático de los artiodáctilos, es el más cercano pariente vivo de los cetáceos.
Un hipopótamo, El más pesado y más acuático de los artiodáctilos, es el más cercano pariente vivo de los cetáceos. La ballena azul es un filtrador de más de 33 metros de largo y el mayor animal de todos los tiempos.
La ballena azul es un filtrador de más de 33 metros de largo y el mayor animal de todos los tiempos.

 El cocodrilo de agua salada es el mayor reptil viviente u un peligro para los seres humanos.
El cocodrilo de agua salada es el mayor reptil viviente u un peligro para los seres humanos. El dragón de Komodo, un gigante insular, es el mayor lagarto y posee una peligrosa saliva infecciosa.
El dragón de Komodo, un gigante insular, es el mayor lagarto y posee una peligrosa saliva infecciosa. La anaconda verde, una constrictora acuática, es la serpiente más pesada, con más de 97.5 kg.
La anaconda verde, una constrictora acuática, es la serpiente más pesada, con más de 97.5 kg.
 La perca del Nilo, uno de los mayores peces de agua dulce, es una peligrosa especie invasora.
La perca del Nilo, uno de los mayores peces de agua dulce, es una peligrosa especie invasora. El tiburón ballena es el pez de mayor tamaño existente, creciendo hasta más de 12.6 metros de largo.
El tiburón ballena es el pez de mayor tamaño existente, creciendo hasta más de 12.6 metros de largo. La manta raya, una especie filtradora, es la mayor de las rayas con más de 7.6 metros, aunque pude remontarse por fuera del agua.
La manta raya, una especie filtradora, es la mayor de las rayas con más de 7.6 metros, aunque pude remontarse por fuera del agua. Examen de un calamar gigante de 9 metros, un gigante abisal y el segundo mayor cefalópodo.
Examen de un calamar gigante de 9 metros, un gigante abisal y el segundo mayor cefalópodo.




![A diferencia del rinoceronte y el mamut lanudos, el buey almizclero logró sobrevivir a las extinciones del Cuaternario.[1]](/wiki/Archivo:Ovibos_moschatus_qtl3.jpg)



.jpg)







{kind=link}
{kind=link}