
Eoceno
| Era[1] | Período | Época | Millones años |
|---|---|---|---|
| Cenozoico | Cuaternario | Holoceno | 0.01 (~10 000 a. C.) |
| Pleistoceno | 2.59 | ||
| Neógeno | Plioceno | 5.33 | |
| Mioceno | 23.03 | ||
| Paleógeno | Oligoceno | 33.90 | |
| Eoceno | 56.00 | ||
| Paleoceno | 66.00 |
El Eoceno es una época y serie de la escala temporal geológica que pertenece al periodo y sistema Paleógeno; dentro de este, el Eoceno sigue al Paleoceno y precede al Oligoceno. Comenzó hace unos 56 millones de años y terminó hace unos 34 millones de años y con una duración de unos 22.1 millones de años.[2][3][4][5]
Durante esta época se comenzaron a formar algunas de las cordilleras más significativas del mundo actual, como los Alpes y el Himalaya, y acontecieron varios cambios climáticos importantes: el máximo térmico del Paleoceno-Eoceno, que aumentó la temperatura del planeta y delimita el inicio de esta época geológica; y el evento Azolla, un enfriamiento global que daría paso a las primeras glaciaciones. La extinción masiva Grande Coupure marca el fin del Eoceno.
Las aves predominaban sobre los demás seres, y los primeros cetáceos comenzaron su desarrollo. Además, la especie de serpiente más grande que ha existido data del Eoceno, y se produjo una gran expansión y diversificación de las hormigas. La Antártida comenzó la época rodeada de bosques tropicales, y lo finalizó con la aparición de los primeros casquetes polares. Existen multitud de yacimientos paleontológicos en diversos lugares del mundo que confirman estos hechos, como el sitio fosilífero de Messel, en Alemania, o la Formación Green River, en Norteamérica.
El nombre de Eoceno, definido por el británico Charles Lyell, proviene de las palabras griegas eos (ἠώς, ‘alba’) y kainos (καινός, ‘nuevo’), haciendo referencia a la aparición de los órdenes modernos de mamíferos durante esta época.
Subdivisiones
[editar]El Eoceno se suele subdividir informalmente en Eoceno inferior (Ypresiense), Eoceno medio (Lutetiano y Bartoniense), y Eoceno superior (Priaboniense). Es menos frecuente subdividirlo en inferior y superior únicamente. En este caso, el Lutetiano pasaría a formar parte del Eoceno inferior, mientras que el Bartoniano haría lo propio en el Eoceno superior.
| Era Eratema |
Periodo Sistema |
Época Serie |
Edad Piso |
Inicio, en millones de años |
|---|---|---|---|---|
| Cenozoico[6] | Cuaternario[6] | |||
| Neógeno | 23,03 | |||
| Paleógeno | Oligoceno | Chattiense | ||
| Rupeliense | ||||
| Eoceno | Priaboniense | |||
| Bartoniense | 41,3 | |||
| Luteciense | 47,8 | |||
| Ypresiense | ||||
| Paleoceno | Thanetiense | |||
| Selandiense | ||||
| Daniense | ||||
- Ypresiense: comenzó hace 55.8 ± 0.2 millones de años, coincidiendo con el inicio del máximo térmico del Paleoceno-Eoceno, un período de calentamiento global rápido e intenso que provocó la extinción de numerosos foraminíferos bentónicos, y que está asociado con un episodio de gran evolución de los mamíferos. En las series estratigráficas, su inicio queda marcado por una variación del isótopo 13C, ya que aumentaron los niveles de CO2 y la relación del isótopo 13C con respecto a 12C disminuyó. El final de esta edad está señalado por un gran desarrollo de los foraminíferos planctónicos y por la aparición del género Hantkenina. Finalizó hace 48.6 ± 0.2 millones de años, y debe su nombre a la localidad de Ypres, en Bélgica.[4]
- Lutetiano: comenzó hace 48.6 ± 0.2 millones de años. Se trata de una edad con abundantes invertebrados marinos (moluscos, corales, erizos de mar) y que se caracteriza por su riqueza en mares epicontinentales sometidos a las influencias continentales. Estratigráficamente, su inicio queda marcado por el desarrollo de los foraminíferos planctónicos y por la aparición del género Hantkenina. Finaliza coincidiendo con la casi-extinción del cocolitóforo Reticulofenestra reticulata, hace unos 40.4 ± 0.2 millones de años, y debe su nombre al antiguo nombre romano de la ciudad de París: Lutetia Parisiorum.[7]
- Bartoniense: comenzó 40.4 ± 0.2 millones de años atrás. Karl Mayer-Eymar le puso nombre, y definió los límites en el año 1857 a partir de sedimentos arcillosos del sur de Inglaterra ricos en fósiles. Estratigráficamente, su inicio queda marcado por la casi-extinción del cocolitóforo Reticulofenestra reticulata. Finalizó hace 37.2 ± 0.2 millones de años, marcado por la primera aparición del cocolitóforo Chiasmolithus oamaruensis. Su nombre proviene de la localidad de Barton-on-Sea, en Inglaterra.[8]
- Priaboniense: Comenzó hace 37.2 ± 0.1 millones de años. Se trata de la última edad del Eoceno, donde tuvo lugar la Grande Coupure, un episodio de extinciones masivas y cambios faunísticos acusados. Estratigráficamente, su inicio queda marcado por la primera aparición del cocolitóforo Chiasmolithus oamaruensis. Finalizó hace 33.9 ± 0.1 millones de años, quedando marcado por los foraminíferos planctónicos y la extinción del género Hantkenina. Su nombre proviene de la localidad de Priabona, en Italia.[5]
Paleogeografía
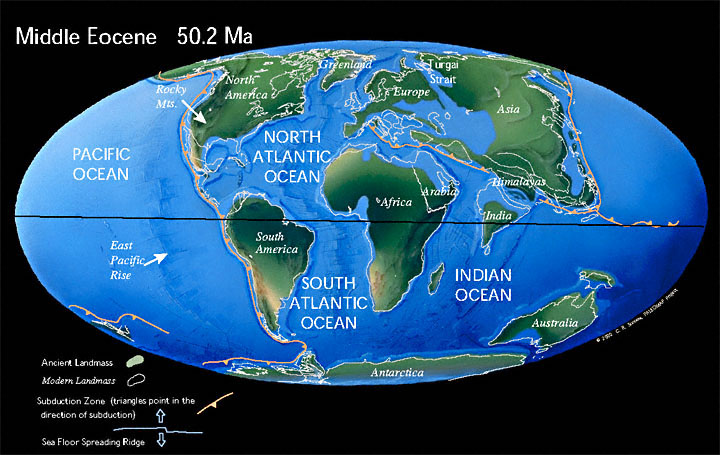
[editar]La tercera y última gran fase de la fragmentación del supercontinente Pangea tuvo lugar a principios del Cenozoico, entre el Paleoceno y el Oligoceno. El paleocontinente Laurentia, formado por los actuales América del Norte y Groenlandia, continuó separándose de Eurasia y ensanchando el joven océano Atlántico, a pesar de que se cree que todavía existía alguna conexión entre ambas masas terrestres.[9] Mientras el Atlántico continuaba su expansión, el ancestral océano Tetis continuó cerrándose debido a la aproximación del continente africano y euroasiático.
A principios del Eoceno, el continente australiano todavía permanecía unido a la Antártida, pero durante el Lutetiano, Australia comenzó a separarse rápidamente de la Antártida moviéndose hacia el norte debido a la deriva continental, tal y como ya había sucedido con el subcontinente Indio y Nueva Zelanda decenas de millones de años antes, durante el Cretácico. El aislamiento del continente antártico acarrearía consecuencias drásticas sobre el clima global, como el máximo térmico del Paleoceno-Eoceno o el evento Azolla.
Orogénesis
[editar]
La Era Cenozoica fue un período de intensa actividad orogénica. Durante la denominada orogenia alpina se formaron las montañas del sistema de Tetis, una cordillera que se extiende sobre la parte meridional de Eurasia y que incluye los Alpes, los Cárpatos, las montañas de Asia menor, Irán, el Hindu Kush, el Himalaya, y las montañas del sureste asiático. Esta actividad orogénica provocó un intenso metamorfismo regional.

El subcontinente indio, que se había separado previamente de Gondwana en el Cretácico Superior, y que se había desplazado a una velocidad de 16 cm/año desde entonces, colisionó con Eurasia a principios del Eoceno. La colisión entre estas dos masas terrestres originó la cordillera más alta del mundo, el Himalaya.[10] Este proceso de orogénesis todavía perdura en la actualidad, haciendo que el Himalaya sea unos cinco centímetros más alto cada año.[11]
Por otra parte, la orogenia cimmeriana, un proceso orogénico que había comenzado en el Jurásico, continuó creando algunas de las cordilleras que actualmente se encuentran en el centro del continente asiático. El Eoceno fue el escenario de la fase final de esta orogénesis.
Eurasia no fue el único continente con actividad orogénica. La configuración geológica de multitud de montañas de América del Norte datan de principios de Cenozoico, como por ejemplo las Black Hills de Dakota del Sur, Wyoming, o las Apalaches de la costa este.
Clima
[editar]
El clima global del Eoceno fue, probablemente, el más homogéneo del Cenozoico; el gradiente térmico del ecuador a los polos era entonces la mitad que en la actualidad, y las corrientes oceánicas profundas eran excepcionalmente cálidas. Las regiones polares eran mucho más cálidas que hoy en día, con temperaturas similares al actual noroeste de los Estados Unidos. Los bosques templados llegaban hasta los mismos polos, mientras que los climas tropicales lluviosos llegaban hasta los 45° de latitud norte. La diferencia más elevada se encontraba en las latitudes templadas, aunque el clima de los trópicos era similar al de nuestros tiempos.[12] Al estar unidos al inicio del Eoceno el continente australiano y la Antártida en una sola masa terrestre, las corrientes oceánicas frías y cálidas se mezclaban, manteniendo una temperatura oceánica homogénea.[13]
Desde el principio de esta época, la temperatura aumentó, en uno de los calentamientos globales más rápidos (en términos geológicos) y extremos que se han registrado en la historia geológica, denominado máximo térmico del Paleoceno-Eoceno. Fue un episodio de calentamiento rápido e intenso (de hasta 7 °C en latitudes altas) que duró menos de cien mil años.[14] El máximo térmico provocó una extinción masiva, por lo que la fauna del Eoceno y del Paleoceno son muy diferentes.
Es posible que este intenso calentamiento fuera causado por la expulsión de clatratos de metano enterrados en el fondo marino. Se cree que los sedimentos donde se encontraban almacenados estos clatratos fueron perturbados a medida que los océanos se calentaban, y el metano que contenían provocó el aumento de la temperatura global. Se estima que se emitieron a la atmósfera dos mil gigatoneladas de metano, un gas de efecto invernadero diez veces más potente que el dióxido de carbono.[15]

El aumento de las temperaturas en todo el planeta no fue la única consecuencia, pues el clima global también se volvió más húmedo, y gran parte de esta humedad fue conducida a los polos.[16] La gran cantidad de lluvia sobre el océano Ártico, junto con la configuración de los continentes (que lo aislaba del resto de océanos), redujo drásticamente la salinidad. El agua dulce acumulada en la zona ártica preparó el terreno para que sucediera un gran cambio climático de signo totalmente opuesto y que marcaría el final del Eoceno.
El clima se mantuvo cálido durante el resto del Eoceno, a pesar de que un enfriamiento global, que finalmente llevaría a las glaciaciones del Pleistoceno, comenzó a mediados de esta época a causa de dos factores: el evento Azolla, y el aislamiento de la Antártida.
El evento Azolla tuvo lugar hace cuarenta y nueve millones de años, cuando el helecho Azolla de agua dulce, también llamado «helecho mosquito», comenzó a crecer en grandes cantidades sobre el océano Ártico. A medida que se hundían sobre suelo marino, las plantas empezaron a formar parte de los sedimentos del suelo oceánico, donde no era posible su descomposición debido al escaso nivel de oxígeno de las capas de agua profundas. La reducción de la cantidad de carbono en la atmósfera terrestre contribuyó a transformar el planeta de una «Tierra invernadero», suficientemente cálida como para que las tortugas y las palmeras habitaran en los polos, en una «Tierra helada».[17][18]
Con la separación del continente australiano hace unos cuarenta y cinco millones de años, la Antártida quedó privada del flujo de aguas ecuatoriales que hasta entonces suavizaba el clima. Con la privación de estas aguas cálidas, la Antártida se enfrió y el océano Antártico inició su glaciación, creando un flujo de agua fría y banquisas que reforzaron el efecto del enfriamiento.
Fauna
[editar]Aves
[editar]Por primera vez en la historia de la Tierra, las aves predominaban sobre todos los demás seres. Las aves predadoras gigantes, como es el caso del Gastornis, anteriormente conocido como Diatryma, se alimentaban de mamíferos como el Propalaeotherium, en Europa y América del Norte,[19] mientras que los Phorusrhacidae, conocidos como las «aves del terror», se convertirían en los superpredadores por excelencia de América del Sur.
Los pingüinos, que habían aparecido durante el Paleoceno, llegaron a América del Sur sobre el Eoceno medio, y en el Bartoniense ya habían comenzado a extenderse por aguas atlánticas. Algunos géneros y especies primitivas de pingüinos son los Perudyptes devriesi (de 76 cm de altura) y el gran Icadyptes salasi (de 150 cm de altura), ambos hallados en las costas desérticas de Ica, al sur del Perú;[20] asimismo los Archaeospheniscus o el gigantesco Anthropornis nordenskjoeldi, que medía 170 cm de altura y pesaba alrededor de 90 kg. En comparación, el pingüino emperador, que es el pingüino actual más grande, solamente mide 122 cm de altura y 37 kg de peso.



Los anseriformes comenzaron a diversificarse, en géneros como Presbyornis, que estaba relacionado con los patos y las ocas actuales. Los estudios de la familia Presbyornithidae son muy importantes para entender la evolución de las aves. Las especies que vivían en zonas costeras y estaban menos especializadas tendían menos a extinguirse que las especies más adaptadas a un ecosistema concreto.
El Palaeotis es una ratite de la cual se han hallado fósiles en el sitio fosilífero de Messel, en Alemania. Estos hallazgos son especialmente interesantes, pues la tesis tradicional sostiene que las ratites se originaron en Gondwana, basándose en su difusión actual. El descubrimiento en Europa de Palaeotis, junto con otras ratites en América del Norte y Mongolia, pone en evidencia esta hipótesis.[21]
En las fosforitas de Quercy se han encontrado evidencias de una difusión más amplia de los loros durante el Eoceno. Los restos fósiles del género Quercypsitta, datados entre 34 y 37 millones de años, indican que los loros llegaron en el pasado a latitudes más septentrionales de las que se encuentran en la actualidad.[22] El fósil de loro más antiguo del que se tiene constancia ha sido hallado en la Formación Fur, Dinamarca, y tiene una antigüedad de 54 millones de años, coincidiendo con el inicio del Eoceno.[23]
Mamíferos
[editar]El acontecimiento más importante en la evolución de los mamíferos durante el Eoceno fue, probablemente, la evolución de los cetáceos. Después de que sus antepasados abandonaran la vida acuática 300 millones de años atrás, un grupo de mamíferos relacionado con los artiodáctilos primitivos consiguió efectuar la transición de un medio terrestre a un medio acuático.
Este proceso comenzó con los Pakicetidae del Eoceno medio e inferior de Pakistán. Se trataba de animales carnívoros terrestres, pero la configuración de los huesos de los oídos y su dentición demostró que representan el primer paso en la evolución de las ballenas. Unos cuantos millones de años más tarde, algunas criaturas, como el Ambulocetus, ya tenían un estilo de vida anfibio, y sus patas posteriores estaban más adaptadas para la natación que para caminar sobre tierra firme.[24] Los Protocetidae representan un paso posterior en la evolución de los cetáceos, y es posible que por aquel entonces ya dispusieran de una aleta caudal como la de los cetáceos actuales.[25]
Los primeros cetáceos completamente marinos aparecieron hace unos cuarenta y cinco millones de años. Los Basilosauridae, que incluyen géneros como el Basilosaurus o el Dorudon, tenían una anatomía muy similar a la de las ballenas actuales. Aun así, su cerebro se encontraba menos desarrollado y no tenían el melón típico de los odontocetos. Las primeras ballenas dentadas no aparecerían hasta casi finalizado el Eoceno.





Los ungulados también continuaron evolucionando durante el Eoceno. Los artiodáctilos aparecieron a principios de esta época, hace cincuenta y cuatro millones de años, y a finales del Eoceno ya se habían diversificado en los tres subórdenes actuales: Tylopoda (camellos), Suinae (cerdos), y Ruminantia (ovejas, cabras, y vacas). La gran expansión de los Perissodactyla, que los desplazaron hacia hábitats menos prósperos, y la aparición de hierba en el Eoceno, condicionaron el desarrollo del particular aparato digestivo que poseen los artiodáctilos y que más adelante les ayudaría a desbancar a los Perissodactyla como los herbívoros dominantes.[26]
Los carnívoros dominantes durante el Eoceno fueron los creodontos. Los carnívoros suelen tener dos dientes carnasiales, uno molar y otro premolar, pero en cambio, los dientes carnisales de los creodontos eran ambos molares.[27] Los creodontos incluyen algunos de los mamíferos predadores terrestres de mayor tamaño que han existido, como el Andrewsarchus, que llegaba a medir tres metros y medio de longitud, casi dos metros de altura, y pesaba 250 kg.[19] Aun así, su gran tamaño no le bastó para imponerse a largo plazo, pues fueron superados por otros carnívoros y terminaron por extinguirse en el Mioceno. Su extinción se debió a una serie de factores, en primer lugar, su articulación lumbosacra no se encontraba suficientemente evolucionada para correr como el resto de carnívoros, y, además, su condición de plantígrado les hacía menos eficientes a la hora de correr.[28] En segundo lugar, los creodontes tenían una dentición diferente que les hacía carnívoros obligados, es decir, estaban restringidos a comer solamente carne, mientras que los miácidos y la mayoría de carnívoros de la época todavía poseían dientes adaptados para masticar otro tipo de alimentos.[27] El último creodonte que aparece en el registro fósil, el Dissopsalis, se extinguió hace ocho millones de años.
Peces
[editar]
Durante el Eoceno, los tiburones lamniformes (o tiburones rayados) sufrieron una gran diversificación. El tiburón duende es uno de tantos ejemplos de tiburones que aparecieron durante esta época.[29] Una de las especies más destacables fue el Otodus obliquus, un tiburón aparecido en el Paleoceno, que podía alcanzar los nueve metros de longitud y se alimentaba de mamíferos marinos, peces, y otros tiburones.[30] Muchos paleontólogos creen que el O. obliquus es un antepasado del género Charcharocles, y, por lo tanto, que mantiene una estrecha relación con el mayor tiburón depredador que ha existido, el Carcharodon megalodon. Otros, sin embargo, relacionan al O. obliquus con el gran tiburón blanco, aunque el número de paleontólogos que lo creen va disminuyendo, pues cada vez existen más evidencias de su relación con el megalodon.[31]
Otro pez destacable fue el Enchodus, un depredador relacionado con los salmones. El Enchodus tenía una serie de colmillos en la parte anterior de los maxilares superior e inferior y en los huesos palatinos. A pesar de ser un depredador, la gran mayoría de sus fósiles se han hallado dentro de los estómagos de otros depredadores más grandes, como los mosasáuridos, los plesiosaurios, o el ave marina Baptornis.
Reptiles
[editar]Una de las especies de serpientes más grande que se conoce vivió durante el Eoceno. Gigantophis garstini podría haber superado los diez metros de longitud, mientras que las serpientes actuales más grandes, las anacondas, rondan los siete metros de largo. Esta serpiente, que habitó el planeta hace cuarenta millones de años en la zona del actual Egipto, se alimentaba probablemente de proboscídeos basales, los antecesores de los actuales elefantes.[32] Otra serpiente de grandes dimensiones que vivió durante esta época fue Palaeophis, una serpiente marina. Al principio, se calculó que su longitud oscilaba entre los treinta y cuarenta metros, una cifra tremendamente elevada para una serpiente, pero estimaciones más recientes arrojan longitudes aproximadas de nueve metros.[33]
Artrópodos
[editar]
El hecho más relevante del Eoceno en relación con los artrópodos fue la expansión de las hormigas. Durante el Cretácico sólo había unas cuantas especies de hormigas en el antiguo continente de Laurasia que representaban menos de un 1 % del total de los insectos. A finales del Paleoceno comenzó una radiación adaptativa que se prolongó durante el Eoceno y que les situó como los insectos dominantes hasta el fin de la época. Su éxito fue tal, que el 90 % de las especies de hormigas que vivieron durante el Eoceno, perduran todavía.[34]
En el sitio fosilífero de Messel, situado en Alemania, se han hallado fósiles de Formicium, un género de hormiga que incluye un total de cinco especies y se conoce únicamente a partir de restos de machos y reinas. La envergadura de las reinas oscilaba entre trece y quince centímetros, lo que la convierte en la mayor de las hormigas que jamás ha existido. También se han encontrado fósiles de este género en el Reino Unido y en Estados Unidos, pero en estos casos se trata solamente de alas aisladas.
Flora
[editar]
A principios del Eoceno, las altas temperaturas calentaron los océanos y crearon un ambiente húmedo y caluroso, donde se podían encontrar bosques que se extendían de polo a polo. Excepto las regiones desérticas más secas y extremas, la Tierra se encontraba completamente cubierta de bosques.[13]
Los bosques polares gozaban de una gran extensión. Se han hallado fósiles e incluso restos preservados de árboles, como las cupresáceas o el género Metasequoia, en la Isla de Ellesmere, situada en el ártico canadiense. Los restos preservados que se han encontrado no se tratan de fósiles, sino de fragmentos originales que se conservaron en aguas pobres en oxígeno en los bosques pantanosos del Eoceno y que después fueron enterrados antes de que se iniciara su descomposición.[35] También se han encontrado fósiles de árboles subtropicales e incluso tropicales del Eoceno en lugares como Groenlandia o Alaska. Las junglas llegaban hasta latitudes tan septentrionales como el noroeste de los Estados Unidos y Europa.[13]
A principios de esta época, llegaron a crecer palmeras en Alaska y en el norte de Europa, aunque a medida que la época avanzaba, y el planeta se iba enfriando, las palmeras comenzaron a ser menos abundantes. Las Metasequoia se encontraban ampliamente extendidas.
El enfriamiento comenzó a mediados de la época. A finales del Eoceno el interior de los continentes ya había comenzado a desecarse, y en algunas zonas los bosques comenzaban a reducirse considerablemente. La hierba, que acababa de aparecer, se encontraba confinada en las riberas de los ríos y todavía no se había extendido por las sabanas y llanuras.[36]
El enfriamiento terrestre fue acompañado por cambios estacionales. Los árboles caducifolios, que estaban más adaptados a los grandes cambios de temperatura, comenzaron a imponerse sobre las especies perennes tropicales. A finales del Eoceno, los bosques caducifolios cubrían ya vastas regiones en los continentes septentrionales, incluyendo América del Norte, Eurasia y el Ártico, mientras que las junglas solamente lograron resistir en América del Sur, India y Australia.
La Antártida, que comenzó el Eoceno envuelta en bosques templados-subtropicales, se enfrió significativamente a medida que pasaba el tiempo. La flora tropical de temperaturas altas desapareció, y a principios del Oligoceno, el continente antártico ya albergaba bosques caducifolios y grandes regiones de tundra.
Gran Ruptura de Stehlin
[editar]
La Gran Ruptura de Stehlin (en francés Grande Coupure) fue un evento de extinción que supuso un gran cambio en la población de diversos organismos en Europa, siendo los mamíferos uno de los grupos más afectados. El paleontólogo suizo Hans Georg Stehlin acuñó su nombre en 1910,[37] haciendo referencia al cambio drástico de los mamíferos europeos.
Este evento, ocurrido hace 33,9 millones de años,[38] ha servido como criterio para definir el límite entre el Eoceno y el Oligoceno, y está caracterizado por las grandes extinciones y por la especiación alopátrida de especies primitivas aisladas. En Asia sucedió un evento similar, al que se le denominó «Remodelado Mongol».
Una de las causas principales de este hecho parece ser el cierre del estrecho de Turgai, lo que unió Europa y Asia y puso fin al aislamiento paleogeográfico de Europa, permitiendo así migraciones masivas de especies entre ambos continentes. Además, la abertura del pasaje de Drake acentuó la corriente circumpolar antártica, iniciando así un enfriamiento progresivo, lo que dio lugar a la formación de un casquete de hielo en la Antártida,[39] así como a la formación de una capa de agua fría sobre los fondos oceánicos. La formación de casquetes provocó una importante disminución del nivel del mar y acentuó el efecto albedo, reflejando la radiación solar y causando un gran descenso de las temperaturas. La capa de agua fría provocó que muchas de las especies que habitaban en aguas cálidas (como el Basilosaurus y algunos tiburones) perecieran, dando lugar a una fauna muy poco diversificada. El cambio climático que estaba teniendo lugar sería el preámbulo de las primeras glaciaciones polares.
Los mamíferos sufrieron una gran renovación. Los taxones de los mamíferos endémicos europeos fueron sustituidos por inmigrantes asiáticos, extinguiéndose así más de la mitad de los mamíferos europeos, y afectando de este modo al resto de la flora y fauna. Los moluscos también sufrieron una gran renovación, especialmente en la costa oeste de los Estados Unidos y en la llanura costera del norte del golfo de México.[40]
Impactos de meteorito
[editar]Algunas teorías señalan el impacto de bólidos sobre Siberia como el principal responsable de este evento debido a las anomalías encontradas en las trazas de iridio, elemento muy útil para detectar los impactos de meteoritos, supuestamente hace 34 millones de años.[41][42] Inicialmente se propuso como un único evento, pero posteriormente se planteó un patrón de extinción en masa escalonada, siendo cada uno de ellos un impacto distinto de cometa o meteorito.[43][44][45] Sin embargo, este modelo de extinción escalonada fue desechado posteriormente al evidenciarse que los impactos fueron anteriores a las extinciones,[46] confirmándose más tarde gracias al estudio de multitud de cortes y sondeos oceánicos.[47] Estos sondeos han sido datados con mucha precisión mediante los foraminíferos planctónicos, concluyendo que su extinción no fue producida por los impactos.[47][48]
El hallazgo posterior de cuarzos con metamorfismos de choque y espinelas ricas en níquel,[49][50][51] así como el descubrimiento de varios cráteres en Siberia (Cráter Popigai) y Norteamérica, han confirmado que hace aproximadamente 35,6 millones de años tres grandes meteoritos impactaron sobre el planeta.[52] Algunos autores han sugerido que estos impactos aceleraron el enfriamiento global,[53][54] aunque los datos isotópicos no apoyan esta aceleración.[39]
Yacimientos paleontológicos
[editar]Estos son algunos de los yacimientos paleontológicos más destacados de la época Eoceno:
Wadi Al-Hitan
[editar]Wadi Al-Hitan (وادي الحيتان, valle de las ballenas en árabe) es una región del desierto occidental de Egipto, donde se han hallado importantes fósiles de cetáceos primitivos. Estos restos representan uno de los principales registros de la historia de la evolución de las especies: la transición de las ballenas de animales terrestres a animales acuáticos. Los fósiles que se han encontrado en Wadi al-Hitan permiten saber que, durante el Eoceno, lo que hoy en día es el desierto del Sahara por aquel entonces era un mar superficial del océano Tetis.
Los fósiles completos o casi completos de Zeuglodon son los hallazgos más destacados del valle, ganándose de este modo el sobrenombre de «Valle de los Zeuglodon». El Dorudon es otro cetáceo prehistórico del que se han encontrado registros fósiles en Wadi al-Hitan. En el 2005 el valle fue declarado Patrimonio de la Humanidad por la Unesco.
Formación Green River
[editar]
La Formación Green River es una formación geológica que se extiende sobre el norte de Utah, el oeste de Colorado, y el suroeste de Wyoming, en los Estados Unidos. En él se pueden encontrar dos zonas bien diferenciadas de barro limoso muy fino, en los que se conservan una gran diversidad de fósiles completos y detallados. La zona más productiva, la denominada «zona de 18 pulgadas», contiene una gran cantidad de fósiles, entre los que destacan los peces, en una serie de capas laminadas de barro, y representan unos cuatro mil años de depósitos. La segunda zona fosilífera se trata de una zona no laminada de algo menos de dos metros de grosor en la que también se pueden encontrar fósiles muy detallados, aunque su extracción se ve dificultada al no estar dispuesta en láminas separables.[55]
Isla Marambio
[editar]La Isla Marambio, también llamada Isla Seymour, es una de las dieciséis grandes islas que rodean la punta de la península antártica.
El capitán noruego Carl Anton Larsen descubrió varios fósiles en la isla en el año 1882, en su primer viaje a la Isla Marambio, a bordo del barco Jason. Desde entonces, la isla ha sido objeto de cuantiosas investigaciones paleontológicas.
Las investigaciones realizadas en la Isla Marambio han ayudado a la comprensión del enfriamiento durante el Eoceno, un proceso que culminó en el inicio de la glaciación de la Antártida. Estudios del carbonato de diversos puntos del océano Antártico sugieren que, en lugar de tratarse de un enfriamiento monotónico de la temperatura durante esta época, en el Eoceno medio hubo un breve episodio de calentamiento. También se han estudiado multitud de fósiles, como algunos pingüinos extintos, diversos bivalvos, y otros animales y plantas.[56]
Arcilla de Londres
[editar]La Arcilla de Londres es un depósito marino conocido por sus fósiles. Se trata del yacimiento fosilífero más importante del Ypresiense del sur de Inglaterra, y es el único lugar de Europa donde se puede encontrar una enorme variedad de fósiles vegetales del Eoceno inferior.
La arcilla fue depositada en un mar que alcanzaba los doscientos metros de profundidad en su región más oriental. Se han encontrado hasta cinco ciclos de deposición (que representan una transgresión seguida por una reducción de la profundidad del mar), sobre todo en la región occidental, más superficial. Cada ciclo comienza con un material basto (incluyendo concentraciones de sílex redondeado), y finaliza con una arcilla cada vez más arenosa.[57]
Los fósiles hallados de animales incluyen bivalvos, gastrópodos, nautilus, cangrejos, langostas, peces (incluyendo dientes de tiburón), reptiles (sobre todo tortugas), y ejemplares de aves poco comunes. También se han hallado algunos fósiles de mamíferos.
En la Arcilla de Londres son muy abundantes los fósiles de plantas, incluyendo frutos y semillas. Hace poco menos de tres siglos que se empezaron a recoger fósiles vegetales, y se han descrito unas 350 especies. Esto hace que la flora de la Arcilla de Londres sea una de las que más variedad tiene en frutos y semillas fósiles.[58]
Sitio fosilífero de Messel
[editar]
El sitio fosilífero de Messel, situado en Hesse, Alemania, y declarado Patrimonio de la Humanidad en 1995, es una antigua cantera de pizarra bituminosa que contiene la muestra mejor conservada de la fauna y flora del Eoceno medio que se conoce. En la mayoría de yacimientos, encontrar esqueletos parciales supone todo un logro, pero en Messel existen numerosos casos de conservación integral, e incluso en algunos se conserva el pelaje, las plumas, o las marcas de piel de algunas especies. La gran diversidad de especies es también un punto a favor, gracias, en parte, a las erupciones de gas. En el yacimiento se han encontrado:
- Más de diez mil peces fosilizados pertenecientes a diversas especies.
- Miles de insectos tanto acuáticos como terrestres, algunos de los cuales se conserva la coloración.
- Gran cantidad de pequeños mamíferos, incluidos caballos enanos, grandes roedores, primates, zarigüeyas, armadillos, parientes del cerdo hormiguero, y murciélagos.
- Numerosas aves, especialmente especies predadoras.
- Cocodrilos, ranas, tortugas, salamandras, y otros reptiles y anfibios.
- Más de treinta restos de plantas, como hojas de palmera, frutos, polen, y núculas.
Véase también
[editar]Notas y referencias
[editar]- ↑ Los colores corresponden a los códigos RGB aprobados por la Comisión Internacional de Estratigrafía. Disponible en el sitio de la International Commision on Stratigraphy, en «Standard Color Codes for the Geological Time Scale».
- ↑ Global Boundary Stratotype Section and Point (GSSP) of the International Commission of Stratigraphy, Status on 2009.
- ↑ International Stratigraphic Chart, 2008
- ↑ a b Rohde, Robert A. (2005). «Ypresian ICS Stage». GeoWhen Database. International Commission on Stratigraphy. Archivado desde el original el 28 de noviembre de 2015. Consultado el 10 de mayo de 2008.
- ↑ a b Rohde, Robert A. (2005). «Priabonian ICS Stage». GeoWhen Database. International Commission on Stratigraphy. Archivado desde el original el 28 de noviembre de 2015. Consultado el 10 de mayo de 2008.
- ↑ a b Tradicionalmente se han usado Terciario y Cuaternario en lugar del actual Cenozoico, con rango de eratemas o eras, usándose Cenozoico como sinónimo de Terciario y subdividido a su vez en Paleógeno y Neógeno. También se puede encontrar Terciario y Cuaternario como sub-eras dentro del eratema o era Cenozoica. Actualmente, en el 2009, el término Terciario (y la subdivisión correspondiente dentro de Cenozoico) ha dejado de ser recomendado por la Comisión Internacional de Estratigrafía para la escala global, quedando el Cenozoico dividido en los sistemas o periodos Paleógeno, Neógeno y Cuaternario.
- ↑ Rohde, Robert A. (2005). «Lutetian ICS Stage». GeoWhen Database. International Commission on Stratigraphy. Archivado desde el original el 24 de mayo de 2008. Consultado el 10 de mayo de 2008.
- ↑ Rohde, Robert A. (2005). «Bartonian ICS Stage». GeoWhen Database. International Commission on Stratigraphy. Archivado desde el original el 4 de febrero de 2008. Consultado el 10 de mayo de 2008.
- ↑ «Tectonics of the Eocene». UC Museum of Paleontology (en inglés). 1999. Consultado el 18 de mayo de 2008.
- ↑ Bin Zhu, William S. F. Kidd, David B. Rowley, Brian S. Currie, Naseer Shafique (2006). «Age of Initiation of the India-Asia Collision in the East-Central Himalaya» (PDF). Journal of Geology (en inglés) 114. p. 641-643. Archivado desde el original el 2 de diciembre de 2005.
- ↑ Jean-Louis Mugnier, Pascal Leturmy, Gérard Vidal (2004). «Kinetics and Sedimentary Balance of the Subhimalayan Zone, West Nepal». Thrust Tectonics and Hydrocarbon Systems 82. p. 115-130. Archivado desde el original el 18 de abril de 2007. Consultado el 18 de mayo de 2008.
- ↑ Stanley, Steven M. (1999). Earth System History. Nueva York: W.H. Freeman and Company. ISBN 0-7167-2882-6.
- ↑ a b c Agaric (2007). «Earth's Eocene Epoch». Associated Content. Archivado desde el original el 2 de enero de 2013. Consultado el 10 de mayo de 2008.
- ↑ Gavin A. Schmidt, Drew T. Shindell (2003). «Atmospheric composition, radiative forcing, and climate change as a consequence of a massive methane release from gas hydrates». Paleoceanography 18 (1). p. 1004, http://dx.doi.org/10.1029/2002PA000757. Archivado desde el original el 20 de octubre de 2011. Consultado el 10 de mayo de 2008.
- ↑ White, Kasey (2003). «Clues to global warming». UC Santa Cruz. Archivado desde el original el 6 de julio de 2008. Consultado el 10 de mayo de 2008.
- ↑ Pagani, M.;Pedentchouk, N.; Huber, M.; Sluijs, A.; Schouten, S.; Brinkhuis, H.; Sinninghe Damsté, J.S.; Dickens, G.R.; Otros (2006). «Arctic hydrology during global warming at the Palaeocene/Eocene thermal maximum». Nature 443 (7111). p. 598.
- ↑ Macdougall, Doug (2004). Frozen Earth: The Once and Future Story of Ice Ages. University of California Press. ISBN 0-520-24824-4.
- ↑ Mulvaney, Kieran (2001). At the Ends of the Earth: A History of the Polar Regions. Washington, D.C.: Island Press. ISBN 1-55963-908-3.
- ↑ a b Haines, Tim (2001). Walking with Beasts. BBC Books. ISBN 0-563-53763-9.
- ↑ «Pinguinos Gigantes Prehistóricos». ElMundo.es. 26 de junio de 2007. Consultado el 5 de octubre de 2008.
- ↑ Chatterjee, Sankar (1997). The Rise of Birds. Johns Hopkins University Press. ISBN 0-8018-5615-9.
- ↑ Mourier-Chauviré, Cécile (1992). «Une nouvelle famille de Perroquets (Aves, Psittaciformes) dans l'Eocène supérieur des Phosphorites du Quercy». Geobios, Mémoire Spécial 14. p. 169-177.
- ↑ David M.Waterhouse, Bent E.Lindow, Nikita V.Zelenkov, Gareth J.Dyke (2008). «Two new parrots (Psittaciformes) from the lower eocene Fur Formation of Denmark». Palaeontology 51 (3). p. 575-582.
- ↑ Sutera, Raymond (2002). «The Origin of Whales and the Power of Independent Evidence». Reports of the National Center for Science Education 20 (5). p. 33-41.
- ↑ Clementz, Marc T., Goswami, A.; Gingerich, P.; Koch, P. (2006). «Isotopic records from early whales and sea cows: contrasting patterns of ecological transition». Journal of Vertebrate Paleontology 26 (2). p. 355-370.
- ↑ Janis, Christine; Jarman, Peter (1984). The Encyclopedia of Mammals. New York: Facts on File. ISBN 0-87196-871-1.
- ↑ a b MacDonald, David (1993). «The Carnassial Connection». Velvet Claw: A Natural History of the Carnivores. BBC Books. ISBN 0-563-20844-9.
- ↑ William Harmon Norton. «Chapter XXI. The Tertiary» (eBook). The Elements of Geology (en inglés). Globusz Publishing. Archivado desde el original el 9 de marzo de 2008. Consultado el 20 de mayo de 2008.
- ↑ Sepkoski, Jack (2002). «A compendium of fossil marine animal genera (Chondrichthyes entry)». Bulletins of American Paleontology 364. p. 560. Archivado desde el original el 10 de mayo de 2012.
- ↑ Renz, Mark (2002). Megalodon: Hunting the Hunter. p. 26-30, Paleo Press. ISBN 0-9719477-0-8.
- ↑ David Ward, Jim Bourdon (2007). «Carcharocles» (en inglés). Elasmo.com. Consultado el 20 de mayo de 2008.
- ↑ New Scientist (2004). «A giant among snakes». New Scientist (2473). p. 17.
- ↑ Dennis Parmley, Melanie DeVore (2005). «Palaeopheid Snakes from the Late Eocene Hardie Mine Local Fauna of Central Georgia». Southeastern Naturalist 4 (4). p. 703–722.
- ↑ David Grimaldi, Donat Agosti (2000). «A formicine in New Jersey Cretaceous amber (Hymenoptera: Formicidae) and early evolution of the ants». Proceedings of the National Academy of Sciences 97 (25). p. 13678–13683.
- ↑ «Eocene Dawn Redwood Fossil Frond from Cache Creek». The Virtual Fossil Museum (en inglés). Consultado el 20 de mayo de 2008.
- ↑ Thomas R. Van Devender, Mark A. Dimmitt. «Desert Grasses». Arizona-Sonora Desert Museum (en inglés). Consultado el 20 de mayo de 2008.
- ↑ H.G. Stehlin (1910). «Remarques sur les faunules de Mammifères des couches eocenes et oligocenes du Bassin de Paris». Bulletin de la Société Géologique de France 4 (9). p. 488-520.
- ↑ Luterbacher, H.P.; Ali, J.R.; Brinkhuis, H.; Gradstein, F.M.; Hooker, J.J.; Monechi, S.; Ogg, J.G.; Powell, J.; Rol, U.; Sanfi lippo, A; Schmitz, B. (2004). «The Paleogene Period». A Geologic Time Scale 2004. Cambridge University Press, Cambridge, p. 384-408. ISBN 0-521-78142-6.
- ↑ a b Livermore, Roy; Nankivell, Adrian; Eagles, Graeme; Morris, Peter (2005). «Paleogene opening of Drake Passage». Earth and Planetary Science Letters 236 (1-2). p. 459-470.
- ↑ Ivany L.C.; Patterson W.P.; Lohmann K.C. (2000). «Cooler winters as a possible cause of mass extinctions at the Eocene/Oligocene boundary». Nature 407. p. 887-890.
- ↑ Ganapathy, R. (1982). «A Major Meteorite Impact on the Earth 65 Million Years Ago: Evidence from the Cretaceous-Tertiary Boundary Clay». Science 216 (4459). p. 885-886.
- ↑ Álvarez, W., Asaro, F., Michel, H.V., Álvarez, L.W. (1982). «Iridium anomaly approximately synchronous with terminal Eocene extinctions». Science 216 (4548). p. 886-888.
- ↑ Hut, P.; Álvarez, W.; Elder, W.P.; Hansen, T.; Kauffman, E.G.; Keller, G.; Shoemaker, E.M.; Weissman, P.R. (1987). «Comet showers as a cause of mass extinctions». Nature 329. p. 118-126.
- ↑ Keller, G. (1986). «Stepwise mass extinctions and impact events: Late Eocene and early Oligocene». Marine Micropaleontology 13. p. 267-293.
- ↑ Kauffman, E.G. (1988). «The dynamics of marine stepwise mass extinction». Revista Española de Paleontología (Extraordinario). p. 57-71.
- ↑ Keller, G.; D'Hondt, S. L.; Orth, C. J.; Gilmore, J. S.; Oliver, P. Q.; Shoemaker, E. M.; Molina, E. (1987). «Late Eocene impact microspherules - Stratigraphy, age and geochemistry». Meteoritics 22 (Marzo). p. 25-60.
- ↑ a b Molina, E.; Gonzalvo, C.; Keller, G. (1993). «The Eocene-Oligocene planktic foraminiferal transition: extinctions, impacts and hiatuses». Geological Magazine 130 (4). p. 483-499.
- ↑ Gonzalvo, C. y Molina, E. (1992). «Bioestratigrafía y cronoestratigrafía del tránsito Eoceno-Oligoceno en Torre Cardela y Massignano (Italia)». Revista Española de Paleontología 7 (2). p. 109-126.
- ↑ Clymer, Aron K.; Bice, David M.; Montanari, Alessandro (1996). «Shocked quartz from the late Eocene: Impact evidence from Massignano, Italy». Geology 24 (6). p. 483-486.
- ↑ Pierrard, O.; Robin, E.; Rocchia, R.; Montanari, A. (1998). «Extraterrestrial Ni-rich spinel in upper Eocene sediments from Massignano, Italy». Geology 26 (4). p. 307-310.
- ↑ Molina, E.; Gonzalvo, C.; Ortiz, S.; Cruz, L.E. (2006). «Foraminiferal turnover across the Eocene-Oligocene transition at Fuente Caldera, southern Spain: no cause-effect relationship between meteorite impacts and extinctions». Marine Micropaleontology 58. p. 270-286.
- ↑ Poag, C.W.; Mankinen, E.; Norris, R.D. (2003). «Late Eocene Impacts: Geologic Record, Correlation and Paleoenvironmental Consequences». From Greenhouse to Icehouse. Columbia University Press, New York, p. 495-510. ISBN 0-231-12716-2.
- ↑ Wonhof, H.B.; Smit, J.; Brinkhuis, H.; Montanari, A.; Nederbragt, A.J. (2000). «Global cooling accelerated by early late Eocene impacts». Geology 28 (8). p. 687-690.
- ↑ Bodiselitsch, B; Montanari, A.; Koeberl, C.; Coccioni, R. (2004). «Delayed climate cooling in the Late Eocene caused by multiple impacts: high-resolution geochemical studies at Massignano». Earth and Planetary Science Letters 223 (3-4). p. 283-302.
- ↑ Kaytee Brenes, Micah Fleming, Nalini Rao, Lani Smith (1999). «The Green River Formation». Localities of the Eocene (en inglés). Consultado el 21 de mayo de 2008.
- ↑ Ivany, L.C.; Lohmann K.C.; Hasiuk F. (2005). «Middle Eocene Warming on Seymour Island, Antartica: Continental Shelf Paleotemperatures Recorded in Molluscan Carbonates». Salt Lake City Annual Meeting (Octubre). p. 16-19. Archivado desde el original el 21 de agosto de 2008. Consultado el 21 de mayo de 2008.
- ↑ Sumbler M.G. (1996). London and the Thames Valley. British Regional Geology series, British Geological Survey. ISBN 0-11-884522-5.
- ↑ Collinson, M. (1983). Fossil plants of the London Clay. The Palaeontological Association. ISBN 0-901702-26-9.
Bibliografía
[editar]En español
[editar]- López Martínez, Nieves (1998). Geologia y paleontologia del Eoceno de La Pobla de Segur, Lleida. Universitat de Lleida. ISBN 84-89727-60-0.
En inglés
[editar]- Donald R. Prothero (1993). The Eocene-Oligocene Transition: Paradise Lost. Columbia University Press. ISBN 0-231-08091-3.
- Donald R. Prothero, Robert J. Emry (1996). The Terrestrial Eocene-Oligocene Transition in North America. Cambridge University Press. ISBN 0-521-43387-8.
- Marie-Pierre Aubry, William A. Berggren, Marie-Pierre Aubry, Spencer G. Lucas (1998). Late Paleocene-Early Eocene Biotic and Climatic Events in the Marine and Terrestrial Records. Columbia University Press. ISBN 0-231-10238-0.
- Gregg F. Gunnell (2001). Eocene Biodiversity: Unusual Occurrences and Rarely Sampled Habitats. Kluwer Academic Pub. ISBN 0-306-46528-0.
- Elizabeth Nesbitt, Donald R. Prothero, Linda C. Ivany (2003). From Greenhouse to Icehouse: The Marine Eocene-Oligocene Transition. Columbia University Press. ISBN 0-231-12716-2.
En alemán
[editar]- Von Koenigswald, W.; Storch, G.; Habersetzer, J. (1997). Messel. Ein Pompeji der Paläontologie. Jan Thorbecke Verlag, Stuttgart. ISBN 3-7995-9083-8.
Enlaces externos
[editar] Wikimedia Commons alberga una categoría multimedia sobre Eoceno.
Wikimedia Commons alberga una categoría multimedia sobre Eoceno.- Eoceno en la web de Celestia
- Paleomapa del Eoceno
- La vida en el Eoceno (en inglés)
- The Eocene and the Eocene-Oligocene transition (en inglés)
- Wadi Al-Hitan (Whale Valley) en la web del Patrimonio de la Humanidad de la UNESCO (en inglés)
- Formación Green River (en inglés)
- Sitio fosilífero de Messel (en alemán)
{kind=link}