
Archaeopteryx
| Archaeopteryx | ||
|---|---|---|
| Rango temporal: 150 Ma - 148 Ma Jurásico Superior | ||
.jpg) | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Chordata | |
| Clase: | Sauropsida | |
| Superorden: | Dinosauria | |
| Orden: | Saurischia | |
| Suborden: | Theropoda | |
| (sin rango): | Avialae | |
| Familia: | Archaeopterygidae | |
| Género: |
Archaeopteryx Meyer, 1861 [nomen conservandum] | |
| Especies | ||
| Sinonimia | ||
A nivel de género Griphosaurus Wagner, 1862 [nomen rejectum] Griphornis Woodward, 1862 [nomen rejectum] Archaeornis Petronievics, 1917 Jurapteryx Howgate, 1984 ¿Wellnhoferia? Elżanowski, 2001 A nivel de especie Griphornis longicaudatus Woodward, 1862 [nomen rejectum] Griphosaurus longicaudatus Wagner, 1862 [nomen rejectum] Archaeopteryx macrura Owen, 1862 [nomen rejectum] Archaeopteryx siemensii? Dames, 1897 Archaeornis siemensii (Dames, 1897) Archaeopteryx owenii Petronievics, 1917 [nomen rejectum] Archaeopteryx recurva Howgate, 1984 Jurapteryx recurva (Howgate, 1984) Archaeopteryx bavarica Wellnhofer, 1993 Wellnhoferia grandis? Elżanowski, 2001 | ||
Archaeopteryx es un género extinto de aves primitivas, aparentemente con caracteres intermedios entre los dinosaurios emplumados y las aves modernas. El nombre deriva del griego antiguo ἀρχαῖος (archaīos), que significa «antiguo», y πτέρυξ (ptéryx), que significa «pluma» o «ala». En español suele castellanizarse como arqueópterix. Desde el siglo XIX ha sido generalmente aceptado por los paleontólogos, y señalado en las obras de consulta como el ave más antigua conocida (miembro del grupo Avialae).[2] Sin embargo, se han identificado aviales potencialmente más antiguos, incluyendo a Anchiornis, Xiaotingia y Aurornis.[3]
Archaeopteryx vivió en el periodo Jurásico Superior, entre 150 a 148 millones de años, en lo que hoy es el sur de Alemania, durante un momento en que Europa era un archipiélago de islas en un mar tropical cálido poco profundo, mucho más cerca del ecuador de lo que está ahora. Similar en forma a una urraca europea, con individuos más grandes, posiblemente, alcanzasen el tamaño de un cuervo.[4] Archaeopteryx podría crecer a alrededor de 0,5 m (1 pie con 8 pulgadas) de longitud. A pesar de su pequeño tamaño, presentaba amplias alas y la habilidad inferida de volar o planear; Archaeopteryx tiene más en común con otros pequeños dinosaurios del Mesozoico que con las aves modernas. En particular, comparte las siguientes características con las deinonicosaurios (dromeosáuridos y trodóntidos): mandíbulas con dientes afilados, tres dedos con garras, una larga cola huesuda y dedos segundos hiperextensibles ("garra asesina"), plumas (que también sugieren homeotermia) y diversas características del esqueleto.[5][6]
Estas características hacen de Archaeopteryx un candidato claro como forma de transición entre los dinosaurios y las aves.[7][8] Por lo tanto, Archaeopteryx juega un papel importante, no solo en el estudio del origen de las aves, sino también en el estudio de los dinosaurios. Fue nombrado a partir de solo una pluma en 1861.[9] Ese mismo año se anunció el primer espécimen de Archaeopteryx completo. A lo largo de los años se han descubierto fósiles de otros diez ejemplares de Archaeopteryx. A pesar de la variación entre estos ejemplares, la mayoría de los expertos consideran que todos los restos que se han descubierto son pertenecientes a una única especie, aunque esto todavía es objeto de debate.
La mayoría de los once fósiles incluyen impresiones de plumas. Debido a que estas plumas son de una forma avanzada (remeras), estos fósiles son evidencia de que la evolución de las plumas se inició antes del Jurásico Superior.[10] El espécimen tipo de Archaeopteryx fue descubierto tan solo dos años después de que Charles Darwin publicara El origen de las especies. El Archaeopteryx parecía confirmar las teorías de Darwin y se ha convertido en una pieza clave de evidencia para el origen de las aves, debate sobre las formas de transición y la confirmación de la evolución.
Descripción
[editar]
Archaeopteryx vivió durante la edad Titoniense Inferior del período Jurásico, hace aproximadamente 150,8-148,5 millones de años.[11] La mayor parte de los especímenes de Archaeopteryx que se han descubierto provienen de la caliza de Solnhofen en Baviera, sur de Alemania, que es un lagerstätte, una formación geológica conocida por sus fósiles magníficamente detallados.[12]
Archaeopteryx era más o menos del tamaño de un cuervo,[4] con amplias alas que eran redondeadas en los extremos y una larga cola en comparación con la longitud del cuerpo. Podía llegar a un máximo de cincuenta centímetros de longitud, con un peso estimado de 0,8 a 1 kilogramo.[4] Las plumas del Archaeopteryx, aunque menos documentadas que sus otras características, son muy similares en su estructura y diseño a las plumas de aves de hoy en día.[12] Pese a la presencia de numerosos rasgos aviares,[13] no obstante, el Archaeopteryx tenía muchas características de los dinosaurios terópodos. A diferencia de las aves modernas, Archaeopteryx tenía pequeños dientes,[12] así como una larga cola ósea, características que compartía con otros dinosaurios de la época.[14]
Debido a que muestra un gran número de características en común tanto con las aves como con los dinosaurios no avianos, Archaeopteryx ha sido usualmente considerado como un vínculo entre ellos.[12] En la década de 1970, John Ostrom, siguiendo la idea de T. H. Huxley de 1868, afirmó que las aves evolucionaron de los dinosaurios terópodos y Archaeopteryx era una pieza crítica para esta afirmación; tenía ciertos rasgos avianos, como la espoleta, plumas de vuelo, alas y un primer dedo del pie parcialmente puesto al revés, junto a características típicas de los terópodos. Por ejemplo, tiene un largo proceso ascendente en el talón, placas interdentales, un proceso obturador del isquion y largos cheurones en la cola. En particular, Ostrom consideró que Archaeopteryx era notablemente similar a la familia de terópodos Dromaeosauridae.[15][16][17][18][19][20][21][22][23]
Plumaje
[editar]
Los especímenes de Archaeopteryx son notables por sus bien desarrolladas plumas de vuelo. Estas eran marcadamente asimétricas y mostraban la estructura propia de las plumas de vuelo de las aves modernas, con vexilos que le daban estabilidad gracias a la disposición de las barbillas, bárbulas y los barbicelos.[24] Las plumas de la cola eran menos asimétricas, de nuevo en consonancia con la situación de las aves modernas, y también tenían vexilos firmes. El pulgar, sin embargo, no tenía un conjunto separado móvil de plumas (la alula).
El plumaje corporal de Archaeopteryx está menos documentado y solo ha sido investigado con propiedad en el bien preservado espécimen de Berlín. Por tanto, como más de una especie puede estar involucrada, la investigación de las plumas del espécimen de Berlín no necesariamente aplica para el resto de las especies de Archaeopteryx. En ese ejemplar hay una especie de "pantalones" de plumas bien desarrolladas en las patas; algunas de estas plumas parecen tener la estructura básica de plumas de contorno, pero están algo descompuestas (carecen de barbicelos como en las aves ratites).[25] Sin embargo, son parcialmente firmes y por tanto capaces de soportar el vuelo.[26]

Un parche de plumas penáceas se encuentra a lo largo de su espalda, el cual es muy parecido a las plumas de contorno del plumaje corporal de las aves modernas por ser simétrico y firme, aunque no tan rígido como las plumas involucradas en el vuelo. Aparte de esto, los rastros de plumas en el espécimen de Berlín se limitan a una especie de "proto-plumón" muy semejante al hallado en el dinosaurio Sinosauropteryx, siendo descompuesto y peludo, siendo posiblemente más parecido en vida al pelo que a las plumas (aunque no en su estructura microscópica). Estas aparecen en el resto del cuerpo, hasta donde estas estructuras están preservadas y no eliminadas por la preparación, y la parte inferior del cuello.[27]
Por otra parte, no hay indicación de que hubiera plumas en la parte superior del cuello y la cabeza. Aunque es concebible que estas zonas estuvieran desnudas, esto también podría ser un efecto de la preservación. Parece que muchos especímenes de Archaeopteryx quedaron empotrados en sedimento anóxico después de quedar a la deriva por algún tiempo sobre sus espaldas en el mar —la cabeza, el cuello y la cola generalmente se doblan hacia abajo, lo cual sugiere que los especímenes apenas habían empezado a descomponerse cuando se hundieron, y los tendones y músculos se relajaron de tal manera que se logró la forma característica de los especímenes fósiles—.[28] Esto podría significar que la piel ya se había ablandado y aflojado, lo cual es reforzado por el hecho de que en algunos especímenes las plumas de vuelo estaban empezando a despegarse al punto de quedar incrustadas en el sedimento. Por tanto se ha hipotetizado que estos especímenes se movieron por el lecho marino en el agua somera por algún tiempo antes de enterrarse, las plumas de cabeza y del cuello se fueron desprendiendo, mientras que las plumas de la cola que estaban mejor sujetas permanecieron en su sitio.[29]
El origen de las plumas constituye un misterio que Archaepteryx no ayuda a resolver, pues las suyas son completamente modernas; de seguro estas empezaron a desarrollarse antes del Jurásico Superior.[30] Sin embargo, se conocen varios dinosaurios celurosaurianos, estrechamente emparentados con los arqueópterix. Algunos de ellos también presentan plumas (como las descubiertas en Velociraptor en el 2007[31]) y otras fibras de estructura más sencilla o protoplumas. Los dinosaurios emplumados confirman que las plumas se originaron antes que las primeras aves como los arqueópterix. Las plumas, al igual que los pelos y las escamas reptilianas, están formadas por una resistente proteína llamada queratina. Esto ha hecho suponer durante mucho tiempo que la pluma procede de una escama "mellada". Actualmente las pruebas genéticas, paleontológicas y embriológicas favorecen otra explicación distinta que se debe a Prum y Brush:[32] las plumas proceden de estructuras huecas y cilíndricas semejantes a espinas.
Coloración
[editar]
En 2011, el estudiante graduado Ryan Carney y sus colegas llevaron a cabo el primer estudio de los colores de un espécimen de Archaeopteryx.[33] Usando la tecnología del microscopio de barrido y un análisis de fluorescencia de rayos X, el equipo fue capaz de detectar la estructura de los melanosomas en el único ejemplar de la pluma descrita en 1861. La estructura resultante fue entonces comparada con las de 87 especies modernas de aves y se determinó con un alto porcentaje de probabilidad que su color era negro. La pluma estudiada probablemente era una pluma de cobertura, la cual podría haber cubierto parcialmente las plumas primarias en las alas. Aunque el estudio no implica que Archaeopteryx fuera enteramente negro, sugiere que tenía una coloración oscura que incluía su cobertura. Carney señaló que esto es coherente con lo que se sabe de las características de vuelo, ya que los melanosomas negros tienen propiedades estructurales que fortalecen las plumas para el vuelo.[34] Un estudio de 2013 publicado en la revista Journal of Analytical Atomic Spectrometry,[35] mostró nuevos análisis de las plumas de Archaeopteryx que revelaban que el animal puede haber tenido un patrón de coloración complejo basado en plumaje oscuro y claro, con las puntas de sus plumas de vuelo siendo principalmente negro en lugar de que la pluma entera fuera de color negro. Pudo haber sido para la ostentación o para el vuelo, pero permanece en la incertidumbre. El escaneo, realizado con una radiación de sincrotón de una fuente de luz para emitir un haz de rayos X y así identificar varios pigmentos, fue hecho únicamente en unas pocas partes de una pluma, dejando la coloración completa de Archaeopteryx en la especulación.[36]
Paleobiología
[editar]Capacidad de vuelo
[editar]Como en las alas de las aves modernas, las plumas de vuelo de Archaeopteryx eran algo asimétricas y sus plumas caudales eran anchas. Esto implica que las alas y la cola era usadas para generar elevación, aunque no está claro si Archaeopteryx era simplemente un planeador o era capaz de volar activamente. La carencia de un esternón óseo sugiere que no era un volador muy fuerte, aunque los músculos de vuelo pueden haber estado sujetos a la gruesa espoleta en forma de bumerán, a los coracoides en forma de placa, o quizás a un esternón cartilaginoso. La orientación hacia los lados de la articulación del glenoideo (el hombro) entre la escápula, el coracoides y el húmero —en vez de la disposición dorsal que se encuentra en las aves modernas— puede indicar que Archaeopteryx era incapaz de elevar sus alas sobre su espalda, un rasgo necesario para el movimiento ascendente en el aleteo del vuelo activo. De acuerdo con un estudio de Philip Senter publicado en 2006, Archaeopteryx era ciertamente incapaz de volar aleteando como las aves modernas, pero bien pudo haber usado una técnica de planeo con un aleteo exclusivamente descendente.[37]
Las alas de Archaeopteryx eran relativamente grandes, los cual pudo haber resultado en una baja pérdida de la velocidad y un radio de giro reducido. La forma corta y redondeada de las alas pudo haber incrementado el arrastre, pero también pudo haber mejorado la habilidad de Archaeopteryx para desplazarse a través de ambientes llenos de obstáculos como los árboles y arbustos (formas similares se observan en las aves que vuelan en ambientes boscosos o de arbustos como los cuervos y faisanes). La presencia de "alas posteriores", es decir plumas de vuelo asimétricas que partían de las patas parecidas a las vistas en dromeosáuridos como Microraptor, también pudo haber ayudado a la movilidad aérea de Archaeopteryx. El primer estudio detallado de las alas posteriores realizado por Longrich en 2006 sugirió que estas estructuras formaban más del 12 % del total del perfil alar. Esto pudo haber reducido la pérdida de velocidad en más del 6 % y el radio de giro en más del 12 %.[38]
Las plumas de Archaeopteryx eran asimétricas. Esto se ha interpretado como evidencia de que era un animal volador, ya que las aves no voladoras tienden a tener plumas simétricas, pero algunos científicos, incluyendo a Thomson y Speakman, han cuestionado esto. Ellos estudiaron más de 70 familias de aves actuales, y encontraron que algunas especies no voladoras tienen cierto rango se asimetría en sus plumas, y que las plumas de Archaeopteryx caen en este rango.[39] No obstante, el grado de asimetría de Archaeopteryx es más típico de los voladores lentos que de las aves no voladoras.[40]

En 2010, Robert L. Nudds y Gareth J. Dyke publicaron en la revista Science un artículo en el cual analizaron los raquis de las plumas primarias de Confuciusornis y Archaeopteryx. El análisis sugirió que los raquis de estos dos géneros eran más delgados y débiles que los de las aves modernas en relación con su peso corporal. Los autores determinaron que Archaeopteryx y Confuciusornis eran incapaces de volar con aleteo.[41] Este estudio fue criticado por Philip J. Currie y Luis Chiappe. Chiappe sugirió que es dificultoso medir los raquis de las plumas fosilizadas, y Currie especuló que Archaeopteryx y Confuciusornis deben de haber sido capaces de volar en algún grado, ya que sus fósiles son preservados en lo que se cree son sedimentos marinos o lacustres, por lo que debieron de haber podido volar sobre las aguas abiertas.[42] Gregory S. Paul también manifestó su desacuerdo con este estudio, afirmando en una respuesta de 2010 que Nudds y Dyke habían sobrestimado los pesos de estas aves primitivas, y que las estimaciones de peso más exactas permitían el vuelo activo incluso con raquis relativamente estrechos. Nudds y Dyke habían considerado un peso de 250 gramos para el espécimen de Múnich de Archaeopteryx, un ejemplar juvenil, basándose en las estimaciones de peso de los especímenes mayores. Paul señaló que una estimación de peso más razonable para el espécimen de Múnich es de cerca de 140 gramos. Paul también criticó las medidas mismas de los raquis, notando que las plumas de este ejemplar tienen una mala preservación. Nudds y Dyke reportaron un diámetro de 0.75 milímetros para la pluma primaria más larga, lo cual Paul no pudo confirmar usando fotografías. Paul midió algunas de las plumas primarias internas, encontrando raquis de 1.25-1.4 milímetros transversalmente.[43] A pesar de estas críticas, Nudds y Dyke mantuvieron sus conclusiones originales. Ellos señalaron que la afirmación de Paul, que un Archaeopteryx adulto debió haber sido un mejor volador que el espécimen juvenil de Múnich, era dudosa. Esto se debe a que, según su razonamiento, requeriría un raquis incluso más grueso, sin que se haya presentado aún evidencia de este.[44] Otra posibilidad es que no hayan alcanzado el vuelo como tal, sino que en lugar de ello usaran sus alas como ayudas para generar sustentación adicional mientras corría sobre el agua de la misma manera que el lagarto basilisco.[45][46]

En 2004, los científicos analizaron un detallado escaneo TAC del neurocráneo del espécimen de Londres de Archaeopteryx y concluyeron que su cerebro era significativamente mayor que el de muchos dinosaurios, indicando que poseía el tamaño cerebral necesario para volar. La anatomía cerebral general fue reconstruida usando el escaneo. La reconstrucción mostró que las regiones asociadas con la visión abarcaban cerca de un tercio del cerebro. Otras áreas bien desarrolladas involucraban la audición y la coordinación muscular.[47] El escaneo del cráneo también reveló la estructura de su oído interno. Esta se parece más a la de las aves modernas que a la de los reptiles no avianos. Estas características tomadas en conjunto sugieren que Archaeopteryx tenía unos afinados sentidos de la audición, el balance, la percepción espacial y la coordinación necesarios para volar.[48] Archaeopteryx tenía una proporción del telencéfalo con respecto al total del cerebro del 78%, como en las aves modernas, distinto de la condición vista en los dinosaurios no celurosaurios como Carcharodontosaurus o Allosaurus, los cuales tenían una anatomía del cerebro y del oído interno más parecida a la de los cocodrilos.[49]
Archaeopteryx continúa jugando un papel importante en los debates científicos acerca del origen y la evolución de las aves. Algunos científicos lo ven como un animal trepador semiarbóreo, siguiendo la idea de que las aves evolucionaron de formas planeadoras habitantes de los árboles (la hipótesis de "caída de los árboles" para la evolución del vuelo propuesta por O. C. Marsh). Otros científicos ven a Archaeopteryx como un rápido corredor sobre tierra, apoyando la idea de que las aves desarrollaron el vuelo al correr (la hipótesis de "desde el suelo hacia arriba" propuesta por Samuel Wendell Williston). Incluso otros han sugerido que Archaeopteryx puede haber estado en casa tanto en los árboles como en el suelo, como los cuervos modernos, y es esta última idea la que se considera mejor apoyada por las características morfológicas. En conjunto, parece que la especie no estaba particularmente especializada para correr sobre el suelo o para posarse. Un escenario delineado por Elżanowski en 2002 sugirió que Archaeopteryx usaba sus alas mayormente para escapar de los depredadores realizando planeos interrumpidos con breves movimientos descendentes para alcanzar sucesivamente ramas más altas, o alternativamente, para cubrir distancias más largas al planear hacia abajo desde precipicios o copas de los árboles.[29]
Por otra parte, Meseguer y colaboradores han publicado en 2012 los ensayos aerodinámicos realizados con un modelo del ave en un túnel de viento, en los que han comprobado que en Archaeopteryx ya se habían desarrollado dispositivos hipersustentadores análogos a los de las aves actuales. Por un lado han probado que la deflexión de cola disminuye la velocidad necesaria para despegar y el consumo de energía necesario y, por otro, que el primer dedo del ala (el más interno) pudo haber tenido significación aerodinámica y haber funcionado como el precursor estructural del álula —pequeño grupo de plumas que presentan las aves más modernas, que permite el vuelo a baja velocidad, como en aterrizajes y despegues.[50]
Crecimiento
[editar]
Un estudio histológico realizado por Erickson, Norell, Zhongue y otros en 2009 mostró que Archaeopteryx tenía un crecimiento relativamente lento en comparación con las aves modernas, presumiblemente porque las porciones más exteriores de los huesos de Archaeopteryx aparecen pobremente vascularizadas;[4] en vertebrados vivos, los huesos pobremente vascularizados se correlacionan con lentas tasas de crecimiento También se supone que todos los esqueletos conocidos de Archaeopteryx vienen de ejemplares juveniles. Debido a que los huesos de Archaeopteryx no pudieron ser seccionados histológicamente en un análisis esqueletocronológico formal (anillo de crecimiento), Erickson y sus colegas utilizaron huesos vascularizados (porosidad) para estimar la tasa de crecimiento de los huesos. Asumieron que el hueso vascularizado deficiente crece a tasas similares en todas las aves y en particular Archaeopteryx. El hueso pobremente vascularizado de Archaeopteryx podría haber crecido tan lento como ocurre en un ánade real (2,5 micrómetros por día) o tan rápido como el de un avestruz (4,2 micrómetros por día). Usando este rango de tasas de crecimiento óseo, los científicos calcularon cuánto tiempo le tomaría "crecer" a cada espécimen de Archaeopteryx al tamaño observable; pudo haber tomado por lo menos 970 días (había 375 días en un año Jurásico tardío) para alcanzar el tamaño adulto (0,8 a 1 kilogramo). El estudio también encontró que los aviares Jeholornis y Sapeornis crecieron de forma relativamente lenta, como lo hizo el dromeosáurido Mahakala. Sin embargo, los aviares Confuciusornis y Ichthyornis crecieron con relativa rapidez, siguiendo una tendencia de crecimiento similar a la de las aves modernas.[51] Una de las pocas aves modernas que presentan un crecimiento lento es el kiwi no volador, y los autores especularon con que Archaeopteryx y el kiwi tuvieron una tasa metabólica basal similar.[4]
Patrones de actividad diaria
[editar]Comparaciones realizadas entre los anillos escleróticos de Archaeopteryx y aves y reptiles modernos indican que puede haber sido un animal diurno, de manera similar a la de muchas aves actuales.[52]
Paleoecología
[editar]
La riqueza y diversidad de la caliza de Solnhofen en la cual se han hallado todos los especímenes de Archaeopteryx ha arrojado luces sobre cómo difería la Baviera del Jurásico de la actual. Su latitud era similar a la de Florida, aunque el clima era probablemente más seco, como se evidencia por los fósiles de plantas con adaptaciones para condiciones áridas y la carencia de sedimentos terrestres característicos de los ríos. La evidencia de plantas, aunque escasa, incluye cícadas y coníferas, mientras que los animales encontrados incluyen un gran número de insectos, lagartos pequeños, pterosaurios y el dinosaurio Compsognathus.[53]
La excelente preservación de los fósiles de Archaeopteryx y de otros animales terrestres en Solnhofen indica que no viajaron muy lejos antes de su preservación.[54] Los fósiles de Archaeopteryx, por tanto, proceden probablemente de individuos que vivieron en las islas bajas que rodeaban la laguna de Solnhofen, en vez de ser originados a partir de cadáveres que llegaron a la deriva desde muy lejos.
Los esqueletos de Archaeopteryx son considerablemente menos numerosos en los depósitos de Solnhofen que los de pterosaurios, de los cuales se han identificado hasta siete géneros distintos.[55] Los pterosaurios incluyen formas tales como Rhamphorhynchus, perteneciente a la familia Rhamphorhynchidae, el grupo que dominó el nicho ecológico actualmente ocupado por las aves marinas y que se extinguieron al final del Jurásico. Los fósiles de pterosaurios, que también incluyen a Pterodactylus, son lo bastante abundantes como para descartar la idea de que eran errantes de las islas mayores, situadas cincuenta kilómetros al norte.[56]
Las islas que rodeaban la laguna de Solnhofen eran semiáridas y subtropicales, con una larga estación seca y pocas lluvias.[57] El análogo moderno más cercano a las condiciones del antiguo Solnhofen sería la cuenca Orca, al norte del golfo de México, aunque es mucho más profunda que las lagunas de Solnhofen.[58] La flora de estas islas estaba adaptada a condiciones secas y consistía en general en arbustos bajos (alrededor de tres metros de alto).[59] Contrariamente a las reconstrucciones de Archaeopteryx que lo muestran trepando por grandes árboles, estos parecen haber estado casi ausentes de las islas; se han hallado pocos troncos en los sedimentos y el polen fosilizado de árboles está ausente.
La forma de vida de Archaeopteryx es difícil de reconstruir y hay varias teorías al respecto. Algunos investigadores han sugerido que estaba principalmente adaptado a la vida en el suelo,[60] mientras que otros creen que era sobre todo arbóreo.[61] La ausencia de árboles no excluye que Archaeopteryx haya tenido un modo de vida arborícola, ya que varias especies de aves actuales viven exclusivamente en arbustos bajos.
El esqueleto de estos animales, del tamaño de una paloma, se parece básicamente al de un dinosaurio terópodo, presentando rasgos propios de este grupo, tales como una larga cola ósea,[62] presencia de garras en los dedos y de dientes en las mandíbulas. Varios aspectos de la morfología de Archaeopteryx, incluyendo la longitud de sus patas y el alargamiento de sus pies, apuntan a modos de vida tanto arborícola como terrestre; algunos expertos consideran probable que hayan sido generalistas, capaces de alimentarse tanto en los arbustos como sobre el terreno abierto, así como a lo largo de las costas de lagunas.[59] Probablemente cazaban presas pequeñas, atrapándolas con sus mandíbulas o con sus garras, dependiendo del tamaño.
Historia
[editar]
Con los años, han sido encontrados doce especímenes de cuerpos fósiles de Archaeopteryx y una pluma que puede pertenecer a este género. Todos los fósiles proceden de depósitos de calizas, extraídos durante siglos, cerca de Solnhofen, Alemania.[53][63]
.jpg)
El descubrimiento inicial, una única pluma, fue desenterrada en 1860 o 1861 y descrita por Christian Erich Hermann von Meyer. Está conservada actualmente en el Museo de Historia Natural de Berlín. Es generalmente asignada a Archaeopteryx y fue el holotipo inicial, pero si es una pluma de esta especie u otra protoave aún por descubrir; se desconoce. Hay algunos indicios de que ciertamente no es del mismo taxón que la mayoría de los esqueletos (el «típico» A. lithographica).[9]
El primer esqueleto, conocido como espécimen de Londres (BMNH 37001),[64] fue descubierto en 1861 cerca de Langenaltheim, Alemania, y quizás fue dado al médico local Karl Häberlein en pago por sus servicios. Häberlein lo vendió por setecientas £ al Museo de Historia Natural de Londres, donde permanece desde entonces.[53] Aunque le falta la mayor parte de su cabeza y cuello, fue descrito en 1863 por Richard Owen como Archaeopteryx macrura, indicando la posibilidad de que no pertenezca a la misma especie que la pluma. Al año siguiente, en la cuarta edición de su obra El origen de las especies,[65] Charles Darwin describió cómo algunos autores sostenían «que la entera clase de las aves arribó a su existencia durante el período Eoceno; pero ahora sabemos, con la autoridad del profesor Owen, que un ave ciertamente vivió durante la deposición de la arenisca verde superior; y aun más recientemente, esta extraña ave, el Archeopteryx, con una larga cola de lagarto, lleva un par de plumas en cada articulación y sus alas decoradas con dos garras libres, ha sido descubierta en las pizarras oolíticas de Solnhofen. Difícilmente algún descubrimiento reciente muestra más vigorosamente que este, cuán poco sabemos aún de los antiguos habitantes del mundo.»[66] Los arqueópterix se convirtieron en una de las tantas pruebas que confirmaron la evolución.[67][68][69]
El término griego "pteryx" (πτέρυξ) se traduce usualmente como "ala", pero también puede significar únicamente "pluma". Von Meyer sugirió esto en su descripción. Al principio él se refirió solo a la pluma que parecía asemejarse a las rémiges (plumas de vuelo) de las aves modernas, pero él había tenido conocimiento del espécimen de Londres gracias a un boceto burdo, al cual se refirió como un «Skelet eines mit ähnlichen Federn bedeckten Thieres» («esqueleto de un animal cubierto de plumas similares»). En alemán, esta ambigüedad se resuelve con el término Schwinge, el cual no alude necesariamente a un ala usada para el vuelo. Urschwinge fue la traducción preferida para Archaeopteryx por los académicos alemanes en el siglo XIX. En español, "pluma de vuelo antigua" ofrece una aproximación cruda a este término.
Desde entonces se han recuperado once especímenes más:
El espécimen de Berlín (HMN 1880) fue descubierto en 1874 o 1875 en Blumenberg, cerca de Eichstätt, Alemania, por el granjero Jakob Niemeyer. Éste vendió el fósil en 1876 al posadero Johann Dörr para poder comprar una vaca, quien a su vez lo revendió a Ernst Otto Häberlein, el hijo de K. Häberlein. Fue puesto a la venta entre 1877 y 1881, lo que atrajo a varios potenciales compradores incluyendo a O. C. Marsh, del Museo Peabody de la Universidad de Yale. Finalmente fue adquirido por 20 000 marcos de oro por el Museo de Historia Natural de Berlín, donde todavía se encuentra conservado. La transacción fue financiada por Ernst Werner von Siemens, fundador de la conocida compañía que lleva su nombre.[53] Descrito en 1884 por Wilhelm Dames, es el espécimen más completo, y el primero con el cráneo completo. En 1897 fue denominado por Dames como una nueva especie, A. siemensii; una revisión reciente apoya la identificación de especie de A. siemensii.[29]
.jpg)
Compuesto únicamente de un torso, el espécimen de Maxberg (S5) fue descubierto en 1956 cerca de Langenaltheim; fue llevado al profesor Florian Heller en 1958, quien lo describió en 1959. El espécimen carece de la cabeza y la cola, aunque el resto del esqueleto está casi completo. Aunque alguna vez fue expuesto en el Museo Maxberg en Solnhofen, actualmente se encuentra en paradero desconocido. Perteneció a Eduard Opitsch, quien lo prestó al museo hasta 1974. Tras su muerte en 1991, se descubrió la pérdida del ejemplar, que bien pudo haber sido robado o vendido.
.jpg)
El espécimen de Haarlem (TM 6428, también conocido como el espécimen de Teyler) fue descubierto en 1855 cerca de Riedenburg, Alemania, y fue descrito como Pterodactylus crassipes en 1857 por von Meyer. Fue reclasificado como Archaeopterix en 1970 por John Ostrom y se encuentra en el Museo Teylers en Haarlem, Países Bajos. Es por tanto el primer espécimen conocido, a pesar del error en su clasificación inicial. También es uno de los especímenes menos completos, consistiendo mayormente en huesos de las extremidades, vértebras cervicales aisladas y algunas costillas. En 2017 fue nombrado un género separado, Ostromia, el cual se considera como más cercanamente relacionado con Anchiornis de China.[70]
El espécimen de Eichstätt (JM 2257) fue descubierto en 1951 cerca de Workerszell, Alemania, y fue descrito por Peter Wellnhofer en 1974. Actualmente se encuentra en el Museo Jura de Eichstätt, Alemania. Es el ejemplar de menor tamaño conocido y el que tiene el segundo cráneo en mejor estado. Podría constituir posiblemente un género separado (Jurapteryx recurva) o al menos una especie separada (A. recurva).
El espécimen de Solnhofen (BSP 1999) fue descubierto en la década de 1970 cerca de Eichstätt y fue descrito en 1988 por Wellnhofer. Está conservado en el Museo Bürgermeister-Müller, en Solnhofen, habiendo siendo clasificado originalmente como Compsognathus por un coleccionista aficionado, el propio burgomaestre Friedrich Müller, por el cual fue bautizado el museo. Es el espécimen de mayor tamaño conocido y puede pertenecer a un género y especie separados, Wellnhoferia grandis. Le faltan algunas partes del cuello, cola, la columna vertebral y el cráneo.
El espécimen de Múnich (S6, anteriormente conocido como el espécimen Solnhofen-Aktien-Verein) fue descubierto el 3 de agosto de 1992 cerca de Langenaltheim y fue descrito en 1993 por Wellnhofer. Se encuentra alojado en el Paläontologisches Museum München, en Múnich, al que fue vendido en 1999 por 1.9 millones de marcos. Posee lo que inicialmente se consideró como un esternón óseo, que más tarde se identificó como parte del coracoides,[71] pero pudo haber tenido un esternón cartilaginoso. Solo le falta el frente del rostro. Podría ser una especie separada, A. bavarica.
.jpg)
Un octavo espécimen fragmentario fue descubierto en 1990, pero no en la caliza de Solnhofen, sino en los sedimentos más recientes de Daiting, en Suabia. Por lo tanto se conoce como el espécimen de Daiting, y había sido conocido desde 1996 a partir de una réplica, brevemente expuesta en el Naturkundemuseum en Bamberg. Permaneció oculto por un largo tiempo, razón por la cual se le apodó el 'fantasma', hasta que el original fue comprado por el paleontólogo Raimund Albertsdörfer en 2009.[72] Estuvo por primera vez expuesto al público junto a otros seis fósiles originales de Archaeopteryx en el Munich Mineral Show en octubre de 2009.[73] Una revisión rápida por los científicos indicó que este espécimen podría ser una nueva especie de Archaeopteryx.[74] Fue hallado en un lecho de caliza que es unos pocos cientos de miles de años más reciente que los demás hallazgos.[72]
.jpg)
Otro fósil fragmentario fue hallado en 2000. Estuvo en posesión de un particular y desde 2004 está en préstamo en el Museo Bürgermeister-Müller de Solnhofen, por lo que se le conoce como el espécimen Bürgermeister-Müller; la propia institución lo designa oficialmente como el «ejemplar de las familias Ottman y Steil, Solnhofen». Como el fragmento representa los restos de una única ala de Archaeopteryx, el nombre popular de este fósil es «ala de pollo».
Por largo tiempo en una colección privada en Suiza, el espécimen de Thermopolis (WDC CSG 100) fue descubierto en Baviera y descrito en 2005 por Mayr, Pohl y Peters. Donado al Wyoming Dinosaur Center en Thermopolis (Wyoming), posee la cabeza y pies mejor preservados; la mayor parte del cuello y la mandíbula inferior no se han preservado. El espécimen de Thermopolis fue descrito en 2 de diciembre de 2005 en un artículo de la revista Science como «Un bien preservado espécimen de Archaeopteryx con rasgos de terópodo»; muestra que Archaeopteryx carecía del dedo del pie trasero «al revés» —un rasgo muy extendido entre las aves— limitando su habilidad de posarse en ramas e implicando que sería un animal terrestre o que treparía solo a los troncos.[75] Esto se ha interpretado como evidencia de su ascendencia terópoda. En 1988, Gregory S. Paul afirmó haber hallado evidencia de un segundo dedo del pie hiperextensible, pero esto no fue verificado y aceptado por otros científicos hasta que el espécimen de Thermopolis fue descrito.[76] «Hasta ahora, este rasgo se pensaba que solo pertenecía a los parientes más cercanos de esta especie, los deinonicosaurios.»[77]
El espécimen de Thermopolis fue asignado a Archaeopteryx siemensii en 2007.[78] Se considera que este espécimen presenta los restos de Archaeopteryx mejor preservados y más completos conocidos.[79]
En 2011 se anunció el descubrimiento de un undécimo espécimen, actualmente conservado en una colección privada. Se dice que es uno de los más completos, pero carece del cráneo y un miembro delantero.[80][81] Su descripción científica fue publicada en julio de 2014 en la revista Nature.[82]
Un duodécimo espécimen fue descubierto por coleccionistas aficionados en 2010 en la cantera Schamhaupten, pero el hallazgo no fue anunciado hasta febrero de 2014.[83] Aún no se ha descrito científicamente.
Clasificación
[editar]
Hasta 2014 los fósiles de este género han sido asignados a una o dos especies, A. lithographica y A. siemensii, pero su historia taxonómica ha sido complicada. Se han publicado docenas de nombres para el manojo de especímenes, muchos de los cuales son simplemente errores de escritura (lapsus). Actualmente se interpreta que el nombre A. lithographica solo se refiere a la única pluma descrita por von Meyer. Sin embargo en 1954 Gavin de Beer concluyó que el espécimen de Londres era el holotipo. En 1960, Swinton propuso que el nombre Archaeopteryx lithographica debía ser situado en la lista oficial de géneros haciendo que los nombres alternativos Griphosaurus y Griphornis fueran inválidos.[84] La ICZN aceptó implícitamente el punto de vista de Beer, llegando a suprimir la plétora de nombres alternativos propuestos en principio para los primeros especímenes de esqueletos,[85] los cuales en su mayoría resultaron de la agria disputa entre von Meyer y su oponente Johann Andreas Wagner (cuyo nombre propuesto Griphosaurus problematicus —«lagarto enigmático problemático»— era una burla mordaz del Archaeopteryx de von Meyer).[86] Adicionalmente, en 1977 se determinó que el primer nombre de especie del espécimen de Haarlem, crassipes, descrito por von Meyer como un pterosaurio antes de que se diera cuenta de su verdadera naturaleza, también debía ser suprimido.[87][88]
Se ha observado que la pluma, el primer espécimen de Archaeopteryx descrito, no se corresponde muy bien con las plumas relacionadas con los vuelos de Archaeopteryx. Ciertamente es una pluma de vuelo de una especie contemporánea, pero su tamaño y proporciones indican que puede pertenecer a otra pequeña especie de terópodo con plumas, de las cuales hasta el momento solo se conoce esta pluma.[9] Como a principios del siglo XXI la pluma fue considerada como el espécimen tipo, esto podría haber creado una significativa confusión de nomenclatura debido a que el nombre Archaeopteryx ya no debería ser aplicado más a los esqueletos. En 2007, dos equipos de científicos realizaron una petición a la ICZN requiriendo que el espécimen de Londres fuera designado explícitamente como el tipo al designarlo como el nuevo espécimen holotipo, o neotipo.[89] Esta sugerencia fue ratificada por la ICZN tras cuatro años de debate, y el espécimen de Londres fue designado como el neotipo en 3 de octubre de 2011.[90]
Especies
[editar]
Si bien se ha argumentado a favor de la idea de que todos los especímenes pertenecen a una misma especie,[91] existen diferencias significativas entre estos. En particular, los especímenes de Múnich, Eichstätt, Solnhofen y Thermopolis se diferencian de los de Londres, Berlín y Haarlem por ser más pequeños o mucho mayores, tener proporciones diferentes de los dedos de la mano, tener hocicos más delgados con dientes apuntando hacia delante, y la posible presencia de un esternón. Debido a estas diferencias, a la mayoría de los especímenes individuales se les ha dado su propio nombre de especie en un momento u otro. El espécimen de Berlín ha sido designado como Archaeornis siemensii, el espécimen de Eichstätt como Jurapteryx recurva, el de Múnich como el Archaeopteryx bavarica, y el de Solnhofen como Wellnhoferia grandis.[29]
En 2007 se publicó una revisión de todos los especímenes bien preservados, incluyendo el entonces recién descubierto espécimen de Thermopolis y llegando a la conclusión de que habría dos especies distintas de Archaeopteryx: A. lithographica (que consta al menos de los especímenes de Londres y Solnhofen), y A. siemensii (que consiste por lo menos en los especímenes de Berlín, Múnich y Thermopolis). Las dos especies se distinguen principalmente por los largos tubérculos flexores en las garras del pie en A. lithographica (las garras de los especímenes de A. siemensii son relativamente más rectas). A. lithographica también tenía una corona más estrecha en algunos dientes y un metatarso más robusto. Una supuesta especie adicional, Wellnhoferia grandis (basada en el espécimen de Solnhofen), parece ser indistinguible de A. lithographica excepto en su tamaño más grande.[79]

A continuación se muestra un cladograma publicado en 2013 por Godefroit et al.[92]
| Avialae |
| |||||||||||||||||||||||||||||||||||||||||||||
Sinónimos
[editar].jpg)

Si se citan dos nombres de autor, el primero denota al descriptor original de la "especie", mientras que el segundo es en quien se basa la combinación binomial dada. Como siempre en la nomenclatura zoológica, colocar el nombre del autor entre paréntesis designa que el taxón fue descrito originalmente en otro género.
- Archaeopteryx lithographica Meyer, 1861 [nomen conservandum]
- Griphosaurus problematicus Wagner, 1862 [nomen oblitum 1961 por opinión 607 de la ICZN]
- Griphornis longicaudatus Woodward, 1862 [nomen oblitum 1961 por opinión 607 de la ICZN]
- Archaeopteryx macrura Owen, 1862 [nomen oblitum 1961 por opinión 607 de la ICZN]
- Archaeopteryx siemensii Dames, 1897
- Archaeornis siemensii (Dames, 1897) Petronievics, 1917[79]
- Archaeopteryx oweni Petronievics, 1917 [nomen oblitum 1961 por opinión 607 de la ICZN]
- Archaeopteryx recurva Howgate, 1984
- Jurapteryx recurva (Howgate, 1984) Howgate, 1985
- Archaeopteryx bavarica Wellnhofer, 1993
- Wellnhoferia grandis Elżanowski, 2001
Los últimos cuatro taxones pueden ser géneros y especies válidas.
"Archaeopteryx" vicensensis (Anon. fide Lambrecht, 1933) es un nomen nudum para lo que parece ser un pterosaurio sin describir.
Controversias
[editar]
Autenticidad
[editar]A principios de 1985, un grupo que incluía al astrónomo Fred Hoyle y al físico Lee Spetner publicaron una serie de artículos en los que se afirmaba que las plumas de los especímenes de Berlín y Londres de Archaeopteryx fueron falsificadas.[93][94][95][96] Sus afirmaciones fueron rechazadas por Alan J. Charig y otros expertos del Museo de Historia Natural de Londres.[97] La mayor parte de su argumentación en favor de una falsificación estaba basada en la falta de familiaridad con los procesos de litificación; por ejemplo, propusieron que, basándose en la diferencia de la textura asociada con las plumas, las impresiones de estas fueron aplicadas sobre una delgada capa de cemento,[98] sin darse cuenta de que las propias plumas podrían haber causado una diferencia de textura.[97] También expresaron su incredulidad ante el hecho de que las losas pudieran dividirse tan suavemente, o que una de las mitades de una losa contuviera fósiles con buena preservación, pero no así la cara opuesta.[99][100] Estas, sin embargo, son propiedades comunes en los fósiles de Solnhofen debido a que los animales muertos pudieron caer en superficies endurecidas que formaban un plano natural para las que las futuras losas se dividieran a lo largo, dejando la mayor parte del fósil en un lado y unos pocos restos en el otro.[97] Ellos también malinterpretaron los fósiles, afirmando que la cola fue falsificada como una sola pluma grande,[98] cuando es visible que ese no es el caso.[97] Adicionalmente, afirmaron que los otros especímenes de Archaeopteryx conocidos en su época no tenían plumas,[99][98] lo cual es incorrecto; los especímenes de Maxberg y Eichstätt tienen plumas obvias.[97] Finalmente, los motivos que sugirieron para la falsificación no son fuertes y son contradictorios; uno es que Richard Owen quería crear evidencia en apoyo de la teoría de la evolución de Charles Darwin, lo cual es improbable dados los puntos de vista de Owen hacia Darwin y su teoría. El otro es que Owen pretendía hacer una trampa a Darwin, esperando que este último apoyara estos fósiles de modo que Owen pudiera desacreditarlo con la falsificación; esto es poco probable debido a que Owen escribió un detallado artículo sobre el espécimen de Londres, por lo que dicha acción podría ciertamente volverse en su contra.[101]
Charig et al. señalaron que la presencia de delgadas fisuras en las losas corriendo a través tanto de la roca como de las impresiones fósiles, así como el crecimiento de mineral sobre las losas que había ocurrido antes de su descubrimiento y preparación son evidencia de que las plumas son genuinas.[97] Entonces Spetner et al. trataron de mostrar que las fisuras pudieron haberse propagado naturalmente a través de la capa de cemento que ellos propusieron,[102] pero negaron considerarlo por el hecho de que las fisuras eran antiguas y habían sido rellenadas con calcita, y por tanto no hubieran sido capaces de propagarse.[103] También intentaron mostrar la presencia de cemento en el espécimen de Londres a través de espectroscopia de rayos X, y ciertamente encontraron algo que no era roca,[102] si bien tampoco era cemento, siendo lo más probable que se tratase de un fragmento de goma de silicona dejado cuando se realizaron moldes del espécimen.[103] Sus sugerencias no han sido tomadas seriamente por los paleontólogos, ya que su evidencia estaba basada mayormente en su falta de conocimiento de la geología, y el hecho de que nunca discutieron sobre los otros especímenes con plumas, los cuales se han incrementado en número desde entonces. Charig et al. reportaron una decoloración: una banda oscura entre las dos capas de caliza que, según ellos, es producto de la sedimentación.[97] Es natural que la caliza tome el color de sus alrededores y muchas calizas están coloreadas (si no con franjas de color) en algún grado, siendo atribuida esta coloración oscura a impurezas.[104] También mencionaron que la completa ausencia de burbujas de aire en las losas de roca es una prueba adicional de que el espécimen es auténtico.[97]
Protoavis y Archaeopteryx
[editar]En 1984, Sankar Chatterjee descubrió fósiles que anunció, para 1991, que pertenecían a un ave extinta mucho más antigua que Archaeopteryx. A estos fósiles, que se cree que datan de entre aproximadamente 210 a 225 millones de años, se les asignó el nombre de Protoavis.[105] Los fósiles estaban tan mal preservados que no permitían estimar si tenía alguna habilidad de vuelo; y aunque las reconstrucciones de Chatterjee usualmente muestran plumas, muchos paleontólogos, incluyendo a Paul (2002) y Witmer (2002), han rechazado las afirmaciones de que Protoavis fuese un ave primitiva (o, incluso, de que en realidad hubiera existido).[59][106] Sus fósiles fueron hallados desarticulados, y fueron recolectados de diferentes localidades. Debido a la deficiente conservación de los restos fósiles, dicho género es considerado por algunos como una quimera, mientras que Archaeopteryx continúa siendo el ave más antigua reconocida universalmente.[107]
Posición filogenética
[editar]
Por lo general, la paleontología moderna clasifica a Archaeopteryx como el ave más primitiva. Sin embargo, no se cree que sea el verdadero ancestro o antepasado de las aves modernas, sino más bien un pariente cercano de dicho ancestro. Aun así, Archaeopteryx ha sido usado frecuentemente como modelo de la verdadera ave ancestral. Varios autores han hecho esto.[106] Lowe (1935)[108] y Thulborn (1984)[109] cuestionaron que Archaeopteryx realmente fue la primera ave. Ellos sugirieron que Archaeopteryx era un dinosaurio que no estaba más cercanamente relacionado con las aves que el resto de los grupos de dinosaurios. Kurzanov (1987) sugirió que Avimimus era un candidato más probable para ser el ancestro de las aves que Archaeopteryx.[110] Barsbold (1983)[111] y Zweers y Van den Berge (1997)[112] señalaron que muchos de los linajes de Maniraptora son extremadamente parecidos a las aves, y por tanto indicaron que los diferentes grupos de aves pueden haber descendido de diferentes ancestros dinosaurios.
El descubrimiento del cercanamente relacionado Xiaotingia en 2011 llevó a nuevos análisis filogenéticos según los cuales Archaeopteryx sería un deinonicosaurio en vez de ser un avial, y por lo tanto, no sería un "ave" bajo los usos más comunes de este término.[2] Un análisis más exhaustivo fue publicado poco después para poner a prueba esta hipótesis, sin poder llegar al mismo resultado; encontraron que Archaeopteryx es un ave en su posición tradicional en la base de Avialae, mientras que Xiaotingia fue encontrado como un deinonicosaurio basal, o un trodóntido. Los autores del estudio siguiente notaron que aún había incertidumbre, y que puede no ser posible establecer con seguridad si Archaeopteryx es un miembro de Avialae o no, salvo que se encuentren nuevos y mejores especímenes de las especies relevantes.[113]
Estudios filogenéticos llevados a cabo por Senter, et al. (2012) y Turner, Makovicky y Norell (2012) confirmaron que Archaeopteryx estaba más cercanamente relacionado con las aves actuales que a los dromeosáuridos y trodóntidos.[114][115] Por otra parte, Godefroit, et al. (2013) encontraron que Archaeopteryx está más emparentado con los dromeosáuridos y trodóntidos en el análisis incluido en su descripción de Eosinopteryx brevipenna. Los autores usaron una versión modificada de la matriz de datos del estudio en que se describió a Xiaotingia, añadiendo a Jinfengopteryx elegans y Eosinopteryx brevipenna a la misma, así como cuatro características adicionales relacionadas con el desarrollo del plumaje. A diferencia del análisis de la descripción de Xiaotingia, los análisis realizados por Godefroit, et al. no encontraron que Archaeopteryx esté particularmente relacionado con Anchiornis y Xiaotingia, los cuales se consideran en cambio como trodóntidos basales.[116]
Agnolín y Novas (2013) encontraron que Archaeopteryx y (su posible sinónimo) Wellnhoferia son los aviales más basales (los autores definieron a Avialae como el grupo que incluye a Archaeopteryx lithographica y Passer, su más reciente ancestro común y todos sus descendientes), junto a Microraptoria, Unenlagiinae, y el clado que contiene a Anchiornis y a Xiaotingia que serían grupos externos sucesivamente relacionados con Avialae.[117] Un estudio filogenético posterior de Godefroit, et al., usando una matriz más inclusiva que aquella de su descripción de Eosinopteryx brevipenna, también encontró que Archaeopteryx es un miembro de Avialae (definido por los autores como el clado más inclusivo que abarca a Passer domesticus, pero no a Dromaeosaurus albertensis o Troodon formosus); se encontró que forma un grado evolutivo en la base de Avialae junto con Xiaotingia, Anchiornis y Aurornis, siendo Xiaotingia un pariente más cercano a las aves modernas y los dos últimos taxones mencionados en una relación más distante con respecto a Archaeopteryx.[118]
En la cultura popular
[editar]Archaeopteryx ha sido considerado por largo tiempo como el ave más primitiva conocida; esto le ha dado una muy extensa atención por parte del público. Su apariencia fácilmente reconocible, y el interés público en los dinosaurios han hecho de Archaeopteryx parte de la cultura popular internacionalmente. Un asteroide del cinturón de asteroides descubierto en 1991, 9860 Archaeopteryx, fue nombrado en honor de este género.[119][120] Una de las apariciones más "extrañas" de Archaeopteryx en la cultura popular se dio en la obra de 1897 de Alfred Jarry, Ubu cocu, ou l'Archéopteryx ('Ubú cornudo, o el Archaeopteryx), que incluye a un Archaeopteryx como un personaje importante.[121]
La película de dibujos animados Fantasía lo incluye en el segmento de «La consagración de la primavera», donde aparece un gran Ornitholestes saltando sobre un pequeño Archaeopteryx.
Galería
[editar]-
 El Archaeopteryx de Londres (1863)
El Archaeopteryx de Londres (1863) -
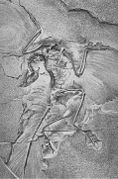 El Archaeopteryx de Berlín (1881)
El Archaeopteryx de Berlín (1881) -
 Representación desactualizada de Archaeopteryx por Heinrich Harder (1916)
Representación desactualizada de Archaeopteryx por Heinrich Harder (1916) -
 Recreación de un Ornitholestes cazando a un Archaeopteryx, de Charles Knight (circa 1910)
Recreación de un Ornitholestes cazando a un Archaeopteryx, de Charles Knight (circa 1910)



)_(17537482124).jpg)
Véase también
[editar] Portal:Dinosaurios. Contenido relacionado con Dinosaurios.
Portal:Dinosaurios. Contenido relacionado con Dinosaurios. Portal:Aves. Contenido relacionado con Aves.
Portal:Aves. Contenido relacionado con Aves.
Referencias
[editar]Notas
[editar]- ↑ Kundrát, Martin; Nudds, John; Kear, Benjamin P.; Lü, Junchang y Ahlberg, Per (2018) «The first specimen of Archaeopteryx from the Upper Jurassic Mörnsheim Formation of Germany». Historical Biology, 31(1): 3-63 doi 10.1080/08912963.2018.1518443
- ↑ a b Xing Xu, Hailu You, Kai Du y Fenglu Han (28 de julio de 2011). «An Archaeopteryx-like theropod from China and the origin of Avialae». Nature 475 (7357): 465-470. PMID 21796204. doi:10.1038/nature10288. Archivado desde el original el 27 de septiembre de 2011.
- ↑ Godefroit, Pascal; Cau, Andrea; Hu, ong-Yu; Escuillié, François; Wu, Wenhao; Dyke, Gareth (2013). «A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds». Nature. en línea. doi:10.1038/nature12168.
- ↑ a b c d e Erickson, Gregory M.; Rauhut, Oliver W. M.; Zhou, Zhonghe; Turner, Alan H.; Inouye, Brian D.; Hu, Dongyu; Norell, Mark A. (2009). «Was Dinosaurian Physiology Inherited by Birds? Reconciling Slow Growth in Archaeopteryx». En Desalle, Robert, ed. PLoS ONE 4 (10): e7390. Bibcode:2009PLoSO...4.7390E. PMC 2756958. PMID 19816582. doi:10.1371/journal.pone.0007390. Archivado desde el original el 4 de abril de 2012. Consultado el 25 de octubre de 2009.
- ↑ Yalden D. W. (1984). «What size was Archaeopteryx?». Zoological Journal of the Linnean Society (en inglés) 82 (1-2): 177-188. doi:10.1111/j.1096-3642.1984.tb00541.x.
- ↑ L. M. Chiappe, L. M. Witme (2002). Mesozoic birds: above the heads of dinosaurs (en inglés). University of California Press: University od California Press Berkeley and Los Angeles, California. p. 151. ISBN 0-520-20094-2.
- ↑ Archaeopteryx: An Early Bird – University of California, Berkeley, Museum of Paleontology. Consultado el 18 de octubre de 2006.
- ↑ Ancient birds flew on all fours Archivado el 14 de julio de 2014 en Wayback Machine. - Nick Longrich, Universidad de Calgary. Discusses how many wings an Archaeopteryx had and other questions.
- ↑ a b c Griffiths, P. J. (1996). «The Isolated Archaeopteryx Feather». Archaeopteryx 14: 1-26.
- ↑ P. Wellnhofer (2004). «The Plumage of Archaeopteryx». En Currie, PJ; Koppelhus, EB; Shugar, MA et al., eds. Feathered Dragons (en inglés). Indiana University Press. pp. 282–300. ISBN 0-253-34373-9.
- ↑ Schweigert, G. (2007). «Ammonite biostratigraphy as a tool for dating Upper Jurassic lithographic limestones from South Germany – first results and open questions». Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen (en inglés) 245 (1): 117-125. doi:10.1127/0077-7749/2007/0245-0117.
- ↑ a b c d Lambert, David (1993). The Ultimate Dinosaur Book (en inglés). Nueva York: Dorling Kindersley. pp. 38-81. ISBN 1-56458-304-X.
- ↑ Holtz, Thomas, Jr. (1995). «Archaeopteryxs Relationship With Modern Birds». Journal of Dinosaur Paleontology (en inglés). Archivado desde el original el 9 de febrero de 2007. Consultado el 1 de marzo de 2007.
- ↑ Palaeogeography, Palaeoclimatology, Palaeoecology 130 (1997) 275-292
- ↑ Bühler, P.; Bock, W. J. (2002). «Zur Archaeopteryx-Nomenklatur: Missverständnisse und Lösung». Journal of Ornithology (Abstract) 143 (3): 269-286. doi:10.1046/j.1439-0361.2002.02006.x. [artículo en alemán, con resumen en inglés]
- ↑ T. H. Huxley (1868). «On the animals which are most nearly intermediate between birds and reptiles.» Geol. Mag. 5, 357-65; Annals & Magazine of Nat Hist 2, 66-75; Scientific Memoirs 3, 3-13.
- ↑ T. H. Huxley (1868) «Remarks upon Archaeopteryx lithographica.» Proc Roy Soc 16, 243-48; Sci Memoirs 3, 340-45.
- ↑ T. H. Huxley (1870) «Further evidence of the affinity between the dinosaurian reptiles and birds.» Quart J Geol Soc 26, 32-50; Sci Mem 3, 487-509.
- ↑ Nedin, C. (1999). All About Archaeopteryx. talk.origins archive. Versión de 10 de junio de 2002. Consultado 18 de octubre, 2006.
- ↑ Olson, S. L. & Feduccia, A. (1979). «Flight capability and the pectoral girdle of Archaeopteryx.» Nature. 278 (5701). 247-248. doi 10.1038/278247a0 (HTML abstract)
- ↑ Ostrom, J. H. (1976). «Archaeopteryx and the origin of birds». Biol. J. Linn. Soc. 8 (2): 91-182. doi:10.1111/j.1095-8312.1976.tb00244.x.
- ↑ Ostrom, J. H. (1985). Introduction to Archaeopteryx. En: Hecht, M. K. O.; Ostrom, J. H.; Viohl, G., & Wellnhofer, P. (eds. [aclaración requerida]) The Beginnings of Birds: Proceedings of the International Archaeopteryx Conference: 9-20. Eichstätt, Freunde des Jura-Museums Eichstätt.
- ↑ Owen, R. (1863). «On the Archaeopteryx of Von Meyer, with a description of the fossil remains of a long-tailed species from the lithographic stone of Solnhofen». Phil. Trans. Roy. Soc. London 153: 33-47. doi:10.1098/rstl.1863.0003.
- ↑ Feduccia, A.; Tordoff, H. B. (1979). «Feathers of Archaeopteryx: Asymmetric vanes indicate aerodynamic function». Science 203 (4384): 1021-1022. Bibcode:1979Sci...203.1021F. PMID 17811125. doi:10.1126/science.203.4384.1021.
- ↑ Chiappe, Luis M. (2007). Glorified Dinosaurs. Sydney: UNSW Press. pp. 118-146. ISBN 0-471-24723-5.
- ↑ Longrich, N. (2006): «Structure and function of hindlimb feathers in Archaeopteryx lithographica.» Paleobiology. 32 (3): 417-431. doi 10.1666/04014.1. (HTML abstract.)
- ↑ Christensen, P; Bonde, N. (2004). «Body plumage in Archaeopteryx: a review, and new evidence from the Berlin specimen». Comptes Rendus Palevol 3 (2): 99-118. doi:10.1016/j.crpv.2003.12.001.
- ↑ Reisdorf, A. G. y M. Wuttke (2012). «Re-evaluating Moodie's Opisthotonic-Posture Hypothesis in fossil vertebrates. Part I: Reptiles - The taphonomy of the bipedal dinosaurs Compsognathus longipes and Juravenator starki from the Solnhofen Archipelago (Jurassic, Germany).» Palaeobiodiversity and Palaeoenvironments; doi 10.1007/s12549-011-0068-y.
- ↑ a b c d Elżanowski A. (2002): «Archaeopterygidae (Upper Jurassic of Germany).» En: Chiappe, L. M., & Witmer, L. M (eds.), Mesozoic Birds: Above the Heads of Dinosaurs: 129-159. University of California Press, Berkeley.
- ↑ P. Wellnhofer (2004). «The Plumage of Archaeopteryx». En Currie PJ, Koppelhus EB, Shugar MA, Wright JL, ed. Feathered Dragons. Indiana University Press. pp. 282-300. ISBN 0-253-34373-9.
- ↑ Turner, A.H.; Makovicky, P. J. y M. A. Norell (2007). «Feather quill knobs in the dinosaur Velociraptor» (pdf). Science 317 (5845): 1721.
- ↑ Prum, R. & A. H. Brush, 2002. «The evolutionary origin and diversification of feathers.» The Quarterly Review of Biology 77: 261-295.
- ↑ a b Carney, R.; Vinther, J.; Shawkey, M. D.; d'Alba, L.; Ackermann, J. (2012). «New evidence on the colour and nature of the isolated Archaeopteryx feather». Nature Communications 3: 637. PMID 22273675. doi:10.1038/ncomms1642.
- ↑ Switek, Brian (9 de noviembre de 2011). «Archaeopteryx was robed in black». New Scientist (Las Vegas).
- ↑ Manning, Phillip. L.; Edwards, Nicholas P.; Wogelius, Roy A.; Bergmann, Uwe; Barden, Holly E.; Larson, Peter L.; Schwarz-Wings, Daniela; Egerton, Victoria M. et al. (2013). «Synchrotron-based chemical imaging reveals plumage patterns in a 150 million year old early bird». Journal of Analytical Atomic Spectrometry 28 (7): 1024. doi:10.1039/c3ja50077b. Archivado desde el original el 17 de junio de 2013.
- ↑ Ghose, Tia (11 de junio de 2013). «Pigments suggest famed dino-bird sported black and white feathers». NBC News. Archivado desde el original el 18 de junio de 2013.
- ↑ Senter, P. (2006). «Scapular orientation in theropods and basal birds and the origin of flapping flight.» Acta Palaeontologica Polonica. 51 (2): 305-313. PDF full text file.
- ↑ Longrich, N. (2006). «Structure and function of hindlimb feathers in Archaeopteryx lithographica». Paleobiology (en inglés) 32 (3): 417-431. doi:10.1666/04014.1.
- ↑ Speakman, J. R.; & Thomson, S. C. (1994). «Flight capabilities of Archaeopteryx». Nature 370 (6490): 514. Bibcode:1994Natur.370..514S. doi:10.1038/370514a0.
- ↑ Norberg, R. A. (1995). «Feather asymmetry in Archaeopteryx». Nature 374 (6519): 211. Bibcode:1995Natur.374..211M. doi:10.1038/374211a0.
- ↑ Nudds, Robert L., & Dyke, Gareth J. (14 de mayo de 2010). «Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability». Science 328 (5980): 887-889. Bibcode:2010Sci...328..887N. PMID 20466930. doi:10.1126/science.1188895.
- ↑ Balter, M. (2010). "Did First Feathers Prevent Early Flight?" Science Now, 2010-5-13.
- ↑ Paul, G. S. (15 de octubre de 2010). «Comment on 'Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability.'». Science 330 (6002): 320. Bibcode:2010Sci...330..320P. doi:10.1126/science.1192963.
- ↑ Dyke, G. J.; Nudds, R. L (15 de octubre de 2010). «Response to Comments on "Narrow Primary Feather Rachises in Confuciusornis and Archaeopteryx Suggest Poor Flight Ability"». Science 330 (6002): 320. Bibcode:2010Sci...330..320N. doi:10.1126/science.1193474.
- ↑ Videler, JJ (2005) Avian Flight. Oxford University Press. ISBN 0-19-856603-4 pages 98-117
- ↑ How Archaeopteryx could run over water (Research Gate article, 27 Dec 2012)
- ↑ Witmer, L. M. (2004). «Palaeontology: Inside the oldest bird brain». Nature 430 (7000): 619-620. Bibcode:2004Natur.430..619W. PMID 15295579. doi:10.1038/430619a.
- ↑ Milner, P. D. (2004). «The avian nature of the brain and inner ear of Archaeopteryx» (PDF). Nature 430 (7000): 666-669. Bibcode:2004Natur.430..666A. PMID 15295597. doi:10.1038/nature02706. Archivado desde el original el 9 de febrero de 2006. Supplementary info
- ↑ Larsson, H. C. E. 2001. Endocranial anatomy of Carcharodontosaurus saharicus (Theropoda: Allosauroidea) and its implications for theropod brain evolution. Pp. 19-33. In: Mesozoic Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- ↑ Meseguer, J.; Chiappe, L. M.; Sanz, J. L.; Ortega, F.; Sanz-Andrés, A.; Pérez-Grande, I. y Franchini, S. (2012) «Lift devices in the flight of Archaeopteryx Archivado el 8 de agosto de 2017 en Wayback Machine.». Spanish Journal of Palaeontology, 27(2): 125-130
- ↑ EurekAlert!, www.eurekalert.org: «Archaeopteryx was not very bird-like.» 8-10-2009.
- ↑ Schmitz, L.; Motani, R. (2011). «Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit Morphology». Science 332 (6030): 705-8. Bibcode:2011Sci...332..705S. PMID 21493820. doi:10.1126/science.1200043.
- ↑ a b c d Chiappe, Luis M. (2007). Glorified Dinosaurs (en inglés). Sydney: UNSW Press. pp. 118-146. ISBN 0-471-24723-5.
- ↑ Davis, P.; and Briggs, D. (1998). «The impact of decay and disarticulation on the preservation of fossil birds». PALAIOS 13 (1): 3-13. JSTOR 3515277. doi:10.2307/3515277. Archivado desde el original el 15 de diciembre de 2007. Consultado el 25 de marzo de 2007.
- ↑ Bartell, K. W.; Swinburne, N. H. M.; Conway-Morris, S. (1994). Solnhofen: a study in Mesozoic palaeontology. Cambridge University Press. ISBN 978-0521458306.
- ↑ Paul, Gregory S. (2002). Dinosaurs of the Air: the Evolution and Loss of Flight in Dinosaurs and Birds. Baltimore: Johns Hopkins University Press. ISBN 0-8018-6763-0.
- ↑ Buisonjé, P. H. de (1985). «Climatological conditions during deposition of the Solnhofen limestones». En Hecht, M. K.; Ostrom, J. H.; Viohl, G. y Wellnhofer, P. (eds. [aclaración requerida]), ed. The Beginnings of Birds: Proceedings of the International Archaeopteryx Conference, Eichstatt, 1984. Eichstätt: Freunde des Jura-Museums Eichstätt. pp. 45-65. ISBN 978-3-9801178-0-7..
- ↑ Bartell, K. W.; Swinburne, N. H. M.; Conway-Morris, S. (1994). Solnhofen: a study in Mesozoic palaeontology (en inglés). Cambridge University Press. ISBN 978-0-521-45830-6.
- ↑ a b c Paul, Gregory S. (2002). Dinosaurs of the Air: the Evolution and Loss of Flight in Dinosaurs and Birds (en inglés). Baltimore: Johns Hopkins University Press. ISBN 0-8018-6763-0.
- ↑ Ostrom, J. H. (1976). «Archaeopteryx and the origin of birds». Biological Journal of the Linnean Society 8 (2): 91-182. doi:10.1111/j.1095-8312.1976.tb00244.x.
- ↑ Feduccia, A. (1993). «Evidence from claw geometry indicating arboreal habits of Archaeopteryx». Science 259 (5096): 790-793. Bibcode:1993Sci...259..790F. PMID 17809342. doi:10.1126/science.259.5096.790.
- ↑ Benton, M. J.; Cook, E.; Grigorescu, D.; Popa, E. y Tallodi, E. (1997). «Dinosaurs and other tetrapods in an Early Cretaceous bauxitefilled fissure, northwestern Romania.» Palaeogeography, Palaeoclimatology, Palaeoecology, 130: 275-292
- ↑ National Geographic News - Earliest Bird Had Feet Like Dinosaur, Fossil Shows - Nicholas Bakalar, 1 de diciembre de 2005, página 1. Consultado el 18 de octubre de 2006.
- ↑ British Museum of Natural History - 'BMNH 37001' - el espécimen tipo
- ↑ Darwin, Origin of Species, Chapter 9, p. 367
- ↑ Darwin, Charles (1859). On the Origin of Species. John Murray. Archivado desde el original el 13 de mayo de 2008. Consultado el 23 de julio de 2013.. Nótese que Darwin escribió 'Archeopteryx', en vez de 'Archaeopteryx'.
- ↑ Esquemas de la composición ósea interna de los arqueopterix. Archivado el 27 de septiembre de 2007 en Wayback Machine. - Restauraciones óseas de de la gran mayoría de los especímenes, realizadas por Jamie Headden, Scott Hartman, y Rutger Jansma's. En este sitio. (ambos obtenidos el 22 de enero de 2007)
- ↑ Archaeopteryx: An Early Bird - Universidad de California Museo de paleontología. Consultado el 18 de octubre de 2006
- ↑ Archaeopteryx lithographica - Nick Longrich, Universidad de Calgary. Discute cuantas alas tienen los arqueópterix, entre otras preguntas.
- ↑ Foth, C.; Rauhut, O.W.M. (2017). «Re-evaluation of the Haarlem Archaeopteryx and the radiation of maniraptoran theropod dinosaurs». BMC Evolutionary Biology 17: 236. doi:10.1186/s12862-017-1076-y.
- ↑ Wellnhofer, P., & Tischlinger, H. (2004). Das "Brustbein" von Archaeopteryx bavarica Wellnhofer 1993 — eine Revision. Archaeopteryx. 22: 3-15. [artículo en alemán].
- ↑ a b Archäologischer Sensationsfund in Daiting, (en alemán) Augsburger Allgemeine - Donauwörth edition; Publicación: 28 de noviembre de 2009; Consultado: 23 de diciembre de 2009.
- ↑ Sammler und Forscher - ein schwieriges Verhältnis (en alemán), Sueddeutsche Zeitung; publicación: 25 de octubre de 2009; Consultado el 25 de diciembre de 2009.
- ↑ Wiedergefundener Archaeopteryx ist wohl neue Art (en alemán). Die Zeit. Consultado: 17 de julio de 2012.
- ↑ Mayr, G; Pohl, B; Peters, DS. (2005). «A well-preserved Archaeopteryx specimen with theropod features». Science 310 (5753): 1483-1486. Bibcode:2005Sci...310.1483M. PMID 16322455. doi:10.1126/science.1120331. Ver comentario en el artículo.
- ↑ Paul, G. S. (1988). Predatory Dinosaurs of the World, a Complete Illustrated Guide. Simon and Schuster, New York. 464 p.
- ↑ National Geographic News - Earliest Bird Had Feet Like Dinosaur, Fossil Shows - Nicholas Bakalar, 1 de diciembre de 2005, página 2. Consultado el 18 de octubre de 2006.
- ↑ Mayr, G.; Phol, B.; Hartman, S.; Peters, D. S. (2007). «The tenth skeletal specimen of Archaeopteryx». Zoological Journal of the Linnean Society 149: 97-116. doi:10.1111/j.1096-3642.2006.00245.x.
- ↑ a b c Mayr, G.; Phol, B.; Hartman, S.; Peters, D. S. (2007). «The tenth skeletal specimen of Archaeopteryx». Zoological Journal of the Linnean Society (en inglés) 149: 97-116. doi:10.1111/j.1096-3642.2006.00245.x.
- ↑ Switek, Brian (19 de octubre de 2011). «Paleontologists unveil the 11th Archaeopteryx». Dinosaur Tracking Blog. The Smithsonian Institution. Archivado desde el original el 22 de octubre de 2011. Consultado el 16 de julio de 2013.
- ↑ Jeff, Hecht (20 de octubre de 2011). «Another stunning Archaeopteryx fossil found in Germany». Short Sharp Science blog. New Scientist. Archivado desde el original el 24 de octubre de 2011.
- ↑ Foth, Christian; Tischlinger, Helmut y Rauhut, Oliver W. M. (2014). «New specimen of Archaeopteryx provides insights into the evolution of pennaceous feathers». Nature 511 (7507): 79-82. doi:10.1038/nature13467.
- ↑ «Copia archivada». Archivado desde el original el 19 de octubre de 2014. Consultado el 13 de junio de 2014.
- ↑ Swinton, W. E. (1960). «Opinion 1084, Proposed addition of the generic name Archaeopteryx VON MEYER, 1861 and the specific name Lithographica, VON MEYER, 1861, as published in the binomen Archaeopteryx Lithographica to the official lists (Class Aves)». Bulletin of Zoological Nomenclature 17 (6-8): 224-226.
- ↑ ICZN (1961). «Opinion 607, Archaeopteryx VON MEYER, 1861 (Aves); Addition to the Official list». Bulletin of Zoological Nomenclature 18 (4): 260-261.
- ↑ A. Wagner (1861). Über ein neues, angeblich mit Vogelfedern versehenes Reptil aus dem Solnhofener lithographischen Schiefer. Sitzungberichte der Bayerischen Akademie der Wissenschaften, mathematisch-physikalische Classe 146-154.
- ↑ Archaeopteryx: An Early Bird - University of California, Berkeley, Museum of Paleontology. Consultado el 18 de octubre de 2006.
- ↑ ICZN (1977). «Opinion 1070. Conservation of Archaeopteryx lithographica VON MEYER 1861 (Aves)». Bulletin of Zoological Nomenclature 33: 165-166.
- ↑ Bock, W. J.; Bühler, P. (2007). «Archaeopteryx lithographica von Meyer, 1861 (Aves): proposed conservation of usage by designation of a neotype». Bulletin of Zoological Nomenclature 64 (4): 261-262.
- ↑ ICZN (2011). «OPINION 2283 (Case 3390) Archaeopteryx lithographica von Meyer, 1861 (Aves): conservation of usage by designation of a neotype». Bulletin of Zoological Nomenclature 68 (3): 230-233.
- ↑ Archaeopteryx turns out to be singular bird of a feather. New Scientist 2443: 17. 17 de abril de 2004. Ver comentarios en el artículo.
- ↑ Godefroit, Pascal; Cau, Andrea; Hu, Dong-Yu; Escuillié, François; Wu, Wenhao; Dyke, Gareth (2013). «A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds.» Nature. doi:10.1038/nature12168. PMID 23719374.
- ↑ Hoyle, F.; Wickramasinghe, N. C. y Watkins, R. S. (1985). «Archaeopteryx». British Journal of Photography 132: 693-694.
- ↑ Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R. y Spetner, L. M. (1985). «Archaeopteryx - a photographic study». British Journal of Photography 132: 264-266.
- ↑ Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R. y Spetner, L. M. (1985). «Archaeopteryx - a further comment». British Journal of Photography 132: 358-359, 367.
- ↑ Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R. y L. M. Spetner, (1985). «Archaeopteryx - more evidence». British Journal of Photography 132: 468-470.
- ↑ a b c d e f g h Charig, A. J.; Greenaway, F.; Milner, A. N.; Walker, C. A. y Whybrow, P. J. (1986). «Archaeopteryx is not a forgery». Science 232 (4750): 622-626. Bibcode:1986Sci...232..622C. PMID 17781413. doi:10.1126/science.232.4750.622.
- ↑ a b c Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R.; Spetner, L. M. (1985). «Archaeopteryx – a photographic study». British Journal of Photography (en inglés) 132: 264-266.
- ↑ a b Hoyle; Wickramasinghe, N. C.; Watkins, R. S. (1985). «Archaeopteryx». British Journal of Photography (en inglés) 132: 693-694.
- ↑ Watkins, R. S.; Hoyle, F.; Wickrmasinghe, N. C.; Watkins, J.; Rabilizirov, R.; Spetner, L. M. (1985). «Archaeopteryx – a further comment». British Journal of Photography (en inglés) 132: 358-359, 367.
- ↑ Nedin, Chris (15 de diciembre de 2007). «On Archaeopteryx, Astronomers, and Forgery». Archivado desde el original el 15 de marzo de 2007. Consultado el 17 de marzo de 2007.
- ↑ a b Spetner, L. M.; Hoyle, F.; Wickramasinghe, N. C. y Magaritz, M. (1988). «Archaeopteryx - more evidence for a forgery». The British Journal of Photography 135: 14-17.
- ↑ a b Nedin, Chris (15 de diciembre de 2007). «On Archaeopteryx, Astronomers, and Forgery» (en inglés). Archivado desde el original el 15 de marzo de 2007. Consultado el 17 de marzo de 2007.
- ↑ https://web.archive.org/web/20091028032011/http://encarta.msn.com/encyclopedia_761565838/Limestone_(mineral).htmlcomo 9 de agosto de 2013. 31 de octubre de 2009.
- ↑ Chatterjee, S. (1991). «Cranial anatomy and relationships of a new Triassic bird from Texas.» Philosophical Transactions of the Royal Society of London, B 332: 277-342.
- ↑ a b Witmer, Lawrence M. (2002). «The debate on avian ancestry». En Witmer, L.; Chiappe, L., ed. Mesozoic Birds: Above the Heads of Dinosaurs. Berkeley: University of California Press. pp. 3-30. ISBN 0-520-20094-2.
- ↑ Ostrom, J. H. (1996). «The questionable validity of Protoavis». Archaeopteryx 14: 39-42.
- ↑ Lowe, P. R. (1935). «On the relationship of the Struthiones to the dinosaurs and to the rest of the avian class, with special reference to the position of Archaeopteryx». Ibis 5 (2): 398-432. doi:10.1111/j.1474-919X.1935.tb02979.x.
- ↑ Thulborn, R. A. (1984). «The avian relationships of Archaeopteryx, and the origin of birds». Zoological Journal of the Linnean Society 82 (1-2): 119-158. doi:10.1111/j.1096-3642.1984.tb00539.x.
- ↑ Kurzanov, S. M. (1987). «Avimimidae and the problem of the origin of birds». Transactions of the joint Soviet-Mongolian Paleontological Expedition 31: 31-94. ISSN 0320-2305.
- ↑ Barsbold, Rhinchen (1983). «Carnivorous dinosaurs from the Cretaceous of Mongolia». Transactions of the joint Soviet-Mongolian Paleontological Expedition 19: 5-119. ISSN 0320-2305.
- ↑ Zweers, G. A.; Van den Berge, J. C. (1997). «Evolutionary patterns of avian trophic diversification». Zoology: Analysis of Complex Systems 100: 25-57. ISSN 0944-2006.
- ↑ Lee, Michael S. Y. y Trevor H. Worthy (2012). «Likelihood reinstates Archaeopteryx as a primitive bird». Biology Letters 8 (2): 299-303. PMC 3297401. PMID 22031726. doi:10.1098/rsbl.2011.0884.
- ↑ Senter, Phil; James I. Kirkland, Donald D. DeBlieux, Scott Madsen y Natalie Toth (2012). «New Dromaeosaurids (Dinosauria: Theropoda) from the Lower Cretaceous of Utah, and the Evolution of the Dromaeosaurid Tail». En Dodson, Peter, ed. PLoS ONE 7 (5): e36790. PMC 3352940. PMID 22615813. doi:10.1371/journal.pone.0036790.
- ↑ Alan Hamilton Turner, Peter J. Makovicky y Mark Norell (2012). «A review of dromaeosaurid systematics and paravian phylogeny». Bulletin of the American Museum of Natural History 371: 1-206. doi:10.1206/748.1.
- ↑ Pascal Godefroit, Helena Demuynck, Gareth Dyke, Dongyu Hu, François Escuillié y Philippe Claeys (2013). «Reduced plumage and flight ability of a new Jurassic paravian theropod from China». Nature Communications 4: Article number 1394. PMID 23340434. doi:10.1038/ncomms2389.
- ↑ Agnolín, Federico L. y Fernando E. Novas (2013). «Avian ancestors. A review of the phylogenetic relationships of the theropods Unenlagiidae, Microraptoria, Anchiornis, and Scansoriopterygidae». SpringerBriefs in Earth System Sciences. SpringerBriefs in Earth System Sciences: 1-96. ISBN 978-94-007-5636-6. doi:10.1007/978-94-007-5637-3.
- ↑ Godefroit, Pascal; Andrea Cau, Hu Dong-Yu, François Escuillié, Wu Wenhao y Gareth Dyke (2013). «A Jurassic avialan dinosaur from China resolves the early phylogenetic history of birds». Nature. in press (7454): 359. doi:10.1038/nature12168.
- ↑ «JPL Small-Body Database Browser: 9860 Archaeopteryx (1991 PW9)». NASA. Consultado el 1 de marzo de 2007.
- ↑ Williams, Gareth. «Minor Planet Names: Alphabetical List». Smithsonian Astrophysical Observatory. Consultado el 1 de marzo de 2007.
- ↑ Buffetaut, E. (1985). «The strangest interpretation of Archaeopteryx». The Beginnings of Birds: Proceedings of the International Archaeopteryx Conference. Eichstätt, Freunde des Jura-Museums Eichstätt. pp. 369-370. ISBN 978-3980117807..
Bibliografía
[editar]- G. R. de Beer (1954). Archaeopteryx lithographica: a study based upon the British Museum specimen. Trustees of the British Museum, Londres.
- P. Chambers (2002). Bones of Contention: The Fossil that Shook Science. John Murray, Londres. ISBN 0-7195-6059-4.
- A. Feduccia (1996). The Origin and Evolution of Birds. Yale University Press, New Haven. ISBN 0-300-06460-8.
- Heilmann, G. (1926). The Origin of Birds. Witherby, Londres.
- T. H. Huxley. (1871). Manual of the anatomy of vertebrate animals. Londres.
- H. von Meyer (1861). Archaeopteryx litographica (Vogel-Feder) und Pterodactylus von Solenhofen. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefakten-Kunde. 1861: 678-679, plate V. [Artículo en alemán]. Texto completo, Google Books.
- P. Shipman (1998). Taking Wing: Archaeopteryx and the Evolution of Bird Flight. Weidenfeld & Nicolson, Londres. ISBN 0-297-84156-4.
- P. Wellnhofer (2008). Archaeopteryx — Der Urvogel von Solnhofen. Verlag Friedrich Pfeil, Munich. ISBN 978-3-89937-076-8.
Enlaces externos
[editar] Wikimedia Commons alberga una galería multimedia sobre Archaeopteryx.
Wikimedia Commons alberga una galería multimedia sobre Archaeopteryx. Wikispecies tiene un artículo sobre Archaeopteryx.
Wikispecies tiene un artículo sobre Archaeopteryx.- Todo sobre Archaeopteryx, de Talkorigins (en inglés)
- Use of SSRL X-ray takes 'transformative glimpse' — Una mirada a los químicos que unen a las aves y a los dinosaurios (en inglés)
- Archaeopteryx: An Early Bird — Universidad de California Museo de Paleontología (en inglés)
- Are Birds Really Dinosaurs? — Universidad de California Museo de Paleontología (en inglés)
| Control de autoridades |
|
|---|
 Datos: Q100196
Datos: Q100196- Multimedia: Archaeopteryx / Q100196
- Especies: Archaeopteryx