
Diferencia entre revisiones de «Acytota»
| Línea 137: | Línea 137: | ||
Actualmente se considera que virus y agentes subvirales evolucionan más rápidamente que los organismos celulares porque que se hacen resistentes a las [[vacuna]]s y su genoma cambia mucho con el paso del tiempo. Los virus y agentes subvirales no tienen [[registro fósil]] debido a que no fosilizan y han pasado inadvertidos durante millones de años. La técnica actual para rastrear el tiempo geológico de los virus es tratar de encontrar [[ADN]] o [[ARN]] viral integrado en los genomas celulares, estas secuencias son llamadas [[Elemento viral endógeno|elementos virales endógenos]],<ref>{{cite journal|last=Feschotte|first=Cédric|autor2=Gilbert, Clement|title=Endogenous viruses: insights into viral evolution and impact on host biology.|journal=Nat Rev Genet.|date=March 2012|volume=13|issue=4|pages=83–96|doi=10.1038/nrg3199}} |
Actualmente se considera que virus y agentes subvirales evolucionan más rápidamente que los organismos celulares porque que se hacen resistentes a las [[vacuna]]s y su genoma cambia mucho con el paso del tiempo. Los virus y agentes subvirales no tienen [[registro fósil]] debido a que no fosilizan y han pasado inadvertidos durante millones de años. La técnica actual para rastrear el tiempo geológico de los virus es tratar de encontrar [[ADN]] o [[ARN]] viral integrado en los genomas celulares, estas secuencias son llamadas [[Elemento viral endógeno|elementos virales endógenos]],<ref>{{cite journal|last=Feschotte|first=Cédric|autor2=Gilbert, Clement|title=Endogenous viruses: insights into viral evolution and impact on host biology.|journal=Nat Rev Genet.|date=March 2012|volume=13|issue=4|pages=83–96|doi=10.1038/nrg3199}} |
||
</ref> ejemplos son los [[ |
</ref> ejemplos son los [[retrovirus endógeno|retrovirus endógenos]] que se derivaron de infecciones virales que atacaron a los primeros mamíferos con antigüedades de más de 180 millones de años y están asociados a la formación de la [[placenta]] en los [[Placentalia|placentarios]].<ref>{{cite journal|last=Taylor|first=D. J.|coauthors=J. Bruenn|title=The evolution of novel fungal genes from non-retroviral RNA viruses|journal=BMC Biology|year=2009|volume=7|doi=10.1186/1741-7007-7-88}}</ref> Otro ejemplo son los [[provirus]] que son secuencias virales que pueden encontrarse en cualquier organismo celular. Estudios de elementos virales endógenos en [[insecto]]s sugieren que los virus ya infectaban a los insectos hace más de 300 millones de años.<ref>https://www.elconfidencial.com/tecnologia/2014-10-19/virus_264212/ </ref> También algunos [[transposones de ADN]] eucariotas como los [[polintones]] y los [[plásmidos]] mitocondriales pudieron haberse originado de infecciones virales que atacaron a los antepasados de los [[eucariotas]] y por tanto podrían interpretarse como fósiles virales. Estas secuencias pueden tener más de 2000 millones de años de acuerdo con la estimación del origen eucariota.<ref>{{cite journal|last1=Krupovic|first1=Mart|last2=Bamford|first2=Dennis H|last3=Koonin|first3=Eugene V|title=Conservation of major and minor jelly-roll capsid proteins in Polinton (Maverick) transposons suggests that they are bona fide viruses|journal=Biology Direct|date=2014|volume=9|issue=1|pages=6|doi=10.1186/1745-6150-9-6|pmid=24773695|pmc=4028283}}</ref><ref>Natalya Yutin, Eugene Koonin, Didier Raoult (2014). [https://pubmed.ncbi.nlm.nih.gov/23701946/ Virophages, Polintons, and Transpovirons: A Complex Evolutionary Network of Diverse Selfish Genetic Elements With Different Reproduction Strategies]. Virology Journal.</ref> |
||
La aparición de las entidades acelulares se estima en el [[Hádico]] hace 4400 millones de años cuando se comenzaron a formar las primeras [[biomolécula]]s.<ref name="RN-20090520"/> |
La aparición de las entidades acelulares se estima en el [[Hádico]] hace 4400 millones de años cuando se comenzaron a formar las primeras [[biomolécula]]s.<ref name="RN-20090520"/> |
||
Revisión del 22:28 4 jul 2020
| Acytota | ||
|---|---|---|
| Rango temporal: 4400–0Ma [1] Hádico - Reciente | ||
 Estructura de algunos tipos de virus. | ||
| Taxonomía | ||
| (sin rango) | Biota | |
| (sin rango) |
Acytota Jeffrey, 1971 | |
| Clasificación de Baltimore | ||
| Grupos, dominios y reinos[2] | ||
| ||
En biología, Acytota (acelular) o Aphanobionta es un grupo taxonómico que describe a los virus y los agentes subvirales en las clasificaciones biológicas.[3][4] Los virus, junto con los agentes subvirales, son clasificados por el Comité Internacional de Taxonomía de Virus. Actualmente dicho comité clasifica a los virus en 4 dominios y 9 reinos.[5]
Actualmente se están construyendo y debatiendo la clasificación de los virus, se han hecho clasificaciones donde se los agrupa dependiendo el tipo de ADN o ARN que porten y el tipo de replicación, a su vez dentro los acelulares existen diversos tipos como los viriones o virus verdaderos y los agentes subvirales que incluye los viroides, virus satélite y virusoides.
Además se cree que varios de sus grupos no están estrechamente emparentados como se piensa y que solo tienen como característica la falta de células rasgo plesiomorfico lo que indicaría que son un taxón parafilético.[6] Los virus poseen proteínas propias que no tienen homólogos con las proteínas celulares,[7] lo que lleva a especular que los acelulares son muy antiguos en la historia de la tierra, especialmente estudios de los proteomas y los pliegues de las proteínas virales que son secuencias que pueden conservarse por miles de millones de años[8] han encontrado que varias familias virales se habrían desarrollado de manera independiente antes que LUCA (último antepasado común universal) siendo las familias de virus de ARN las más antiguas de la naturaleza.[9][10] Otros acelulares como los viroides, virusoides y virus satélite de ARN poseen genes y métodos replicativos muy antiguos, lo que implica que son los restos sobrevivientes de las primeras moléculas autorreplicantes de ARN correspondiente a la Hipótesis del mundo de ARN.[11][12] Varias de estas moléculas se habrían juntado y autoensamblado logrando posteriormente desarrollar una membrana dando origen a las primeras células. Los virus y agentes subvirales también se habían autoensamblado pero no desarrollaron una membrana celular, por lo tanto los virus y agentes subvirales continuaron su ciclo como parásitos intracelulares y con pocos cambios evolutivos.[13][14]
También se puede considerar como acelulares a los plásmidos que son comparables a los virus y agentes subvirales en compartimiento replicativo, transmisión entre huéspedes y por ser autónomos del genoma del huésped. A diferencia de los virus y agentes subvirales que son infectivos los plásmidos son endosimbiontes mutualistas que le confieren beneficios a sus células huésped como la transmisión de genes de resistencia a antibióticos. Los plásmidos están estrechamente emparentados con los virus de ADN según estudios de la ADN polimerasa.
La inclusión de Acytota en el árbol filogenético hace que la vida celular deba definirse en un superdominio Cytota (celular) para diferenciarlos de los virus que son acelulares. Acytota y Cytota son situados en un taxón llamado Biota.
La principal razón que se ha usado para excluir a este grupo del árbol filogenético es la ausencia de ribosomas. Además a todos les falta un ácido nucleico y no tienen registro fósil.
Sinonimia
Se han usado los términos Aphanobionta (Novak 1930, Rothmaler 1951), Archaeophyta (Barkley 1939 non Archephyta-Haeckel 1866), Archetista (Jahn & Jahn 1949), Acellularae (Traub 1963), Protobiota (Hsen-Hsu 1965), Acytota (Jeffrey 1971) y Akamara (Hurst, 2000).
Consideraciones generales

Se ha vuelto a proponer este término para evitar la polémica que causa considerar a los virus y los agentes subvirales como seres vivos, puesto que, aunque contienen material genético propio, todos son capaces de replicación, ninguno de ellos contiene células, es decir, son acelulares (de ahí el término Acytota).
Para que pudieran considerarse seres vivos tendrían que pasar la prueba de la teoría celular, la cual establece que:
- La célula es la unidad básica y estructural de los seres vivos; todos los organismos están compuestos por células.
- Todas se forman por la división de células preexistentes. Cada una contiene material genético que se transmite.
- Toda función química y fisiológica básica ocurre en el interior de la célula.
- Las células contienen información hereditaria indispensable para regular sus funciones y ser transmitida a las siguientes generaciones.
Entre las razones por las que no se considera a este imperio como vivo, además de la prueba de teoría celular, se citan el que necesitan reproducirse dentro de una célula huésped, no pueden realizar ningún intercambio proteínico fuera de una célula hospedadora, ni de ADN y ARN, y que también, por ejemplo, un virus no puede entrar en un huésped sin inducirle una enfermedad o una mutación.
Todos los conformantes de este gran imperio, no conocido en toda su variedad, pueden surgir dentro de un huésped en cualquier momento, por lo que son considerados como entidades virales o subvirales. Además, su tamaño los coloca como los seres ultramicroscópicos, justo a continuación de las proteínas.
Como última razón, después de entrar en un huésped al intentar ser eliminados es casi imposible hasta ahora liberarse totalmente de ellos, ya que aunque sean destruidos, sus genes continúan mostrándose en las células dañadas por estos de por vida.
Tipos de acelulares


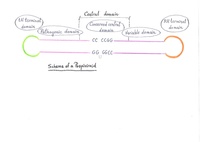

Los acelulares se clasifican dependiendo el tipo de material genético que porten, sin embargo presentan diversas composiciones genómicas o biomoleculares:[15]
- Virión o virus verdadero: Compuestos por material genético puede ser ADN o ARN, una cápside proteica y, en algunos tipos, una envoltura lipídica. Se especializan en infectar cualquier tipo de organismo como los bacteriófagos en bacterias.
- Viroide: Están compuestos únicamente por moléculas de ARN, no poseen cápside proteica y envoltura lipídica. Infectan exclusivamente plantas.
- Virus satélite: Comparten las mismas características que los viriones o viroides, pero se diferencian de estos en que son más pequeños y necesitan un virus auxiliar (virión) para la infección. Por tanto se consideran virus dependientes de otros virus. Dentro de los virus satélite se pueden distinguir algunos subtipos:
- Plásmido: Están compuestos únicamente por moléculas de ADN y viven como endosimbiontes mutualistas dentro los procariotas y en algunos eucariotas como las levaduras. Los plásmidos no son infectivos y no forman parte del genoma del huésped, les pueden conferir beneficios a sus células huéspedes como la transferencia de genes de resistencia a antibióticos.
Actualmente se han descrito más de 6590 especies de acelulares,[16] pero se creé que pueden haber millones de especies que todavía no han sido halladas.[17] La mayor parte de la diversidad acelular se ha descrito exclusivamente en los eucariotas, sin embargo falta por describir más acelulares en los procariotas.
Clasificación de Baltimore
Por otra parte la clasificación de Baltimore ordena a los virus en los siguientes grupos. Esta clasificación también incluye virus satélite y virusoides, pero excluye a los viroides porque sus características replicativas no se adjuntan a la clasificación:[18][19]
- I: Virus ADN bicatenario
- II: Virus ADN monocatenario
- III: Virus ARN bicatenario
- IV: Virus ARN monocatenario positivo
- V: Virus ARN monocatenario negativo
- VI: Virus ARN monocatenario retrotranscrito
- VII: Virus ADN bicatenario retrotranscrito
Origen y evolución

El origen de los acelulares y sus características son controvertidas y difíciles de resolver, para explicar el origen de los acelulares se han propuesto tres hipótesis:[20]
- Hipótesis del virus primero: esta hipótesis considera que los acelulares evolucionaron de complejas moléculas de proteínas y ácidos nucleicos durante el mundo de ARN, antes de la aparición de los organismos celulares y fueron dependiente de los organismos celulares durante toda la historia de la tierra. Esto está respaldado por estudios en los virusoides y los viroides en que se sugiere que son los restos sobrevivientes de las primeras moléculas de ARN autorreplicantes (Hipótesis del mundo de ARN), los cuales pueden representar estadios intermedios en la evolución de los virus. Estudios científicos han revelado que dos tipos de viriones se originaron de manera independiente antes que LUCA (último antepasado común universal).[14] [21]
- Hipótesis de la reducción: esta hipótesis considera que los virus son descendientes de un linaje celular extinto (desconocido) que producto de su parasitismo hayan perdido la estructura celular y ciertos genes que es tal que sus descendientes actuales ya no están conformados por una célula. Bacterias como Rickettsia y Chlamydia son células vivientes que, como los virus, solo pueden reproducirse y activar su metabolismo dentro de células huéspedes.
- Hipótesis del escape: esta hipótesis considera que los virus se originaron desde elementos genéticos móviles (retrotransposoness, transposones). Estos son moléculas de ADN que se multiplican y se mueven a diferentes posiciones en el interior de los genes de la célula. Los seguidores de esta hipótesis incluyen a los elementos genéticos móviles en el dominio Acytota.[6][22]
Ninguna de estas hipótesis se consideraba plenamente aceptada, por ejemplo la hipótesis de la reducción no explicaba porque los virus no se parecen en nada a los parásitos celulares, la hipótesis del escape no explica el origen de las cápsides y las envolturas víricas y la hipótesis del virus primero no explica porque los virus dependen de un huésped celular para su replicación. Un factor que impide identificar a los antepasados virales son la transferencia horizontal de genes y enzimas, ya que los virus y los organismos celulares pueden intercambiar genes y enzimas durante la infección.[23][24] Según las investigaciones científicas las hipótesis más plausibles para el origen de los virus son las del virus primero y la del escape.
El descubrimiento de los acelulares que son fundamentalmente diferentes de los organismos celulares (al ser entidades acelulares) tiene implicaciones en el origen de la vida ya que respalda la hipótesis de que la vida pudo haber comenzado con moléculas orgánicas de autoensamblaje.[13][25][9][10]
Actualmente se considera que virus y agentes subvirales evolucionan más rápidamente que los organismos celulares porque que se hacen resistentes a las vacunas y su genoma cambia mucho con el paso del tiempo. Los virus y agentes subvirales no tienen registro fósil debido a que no fosilizan y han pasado inadvertidos durante millones de años. La técnica actual para rastrear el tiempo geológico de los virus es tratar de encontrar ADN o ARN viral integrado en los genomas celulares, estas secuencias son llamadas elementos virales endógenos,[26] ejemplos son los retrovirus endógenos que se derivaron de infecciones virales que atacaron a los primeros mamíferos con antigüedades de más de 180 millones de años y están asociados a la formación de la placenta en los placentarios.[27] Otro ejemplo son los provirus que son secuencias virales que pueden encontrarse en cualquier organismo celular. Estudios de elementos virales endógenos en insectos sugieren que los virus ya infectaban a los insectos hace más de 300 millones de años.[28] También algunos transposones de ADN eucariotas como los polintones y los plásmidos mitocondriales pudieron haberse originado de infecciones virales que atacaron a los antepasados de los eucariotas y por tanto podrían interpretarse como fósiles virales. Estas secuencias pueden tener más de 2000 millones de años de acuerdo con la estimación del origen eucariota.[29][30]
La aparición de las entidades acelulares se estima en el Hádico hace 4400 millones de años cuando se comenzaron a formar las primeras biomoléculas.[1]
Estudios de las proteínas que codifican ARN polimerasa, ADN polimerasa y la transcriptasa inversa han identificado que los virus pueden dividirse en 4 dominios de manera monofilética. De acuerdo con estos estudios los virus de ARN y los virus retrotranscritos forman un dominio monofilético Riboviria dividido en dos reinos Orthornavirae que incluye los virus de ARN y Pararnavirae que incluye los virus retrotranscritos.[31] A su vez identificaron 3 dominios de virus de ADN, Duplodnaviria, Varidnaviria y Monodnaviria.[32][33][34] En cuanto al origen de estos virus los autores sugirieron que los bacteriófagos de Varidnaviria parecen haber originado superfamilias de transposones de ADN y los plásmidos mitocondriales a través de infecciones virales y a su vez estos transposones y plásmidos parecen haber originado a los adenovirus y los virófagos como consecuencia de una activación o un evento de recombinación genética.[34] Los virus del dominio Monodnaviria pudieron haber evolucionado de plásmidos que codificaban la endonucleasa HUH a través de un evento de recombinación genética que fusionaron el ADN de los plásmidos anteriormente mencionados con las proteínas de la cápside de ciertos virus de ARN.[9][33] Los virus retrotranscritos (Pararnavirae) pudieron haberse originado varias de retrotransposones LTR en eventos de recombinación genética que fusionaron el ácido nucleico de los retrotransposones con las proteínas de ciertos virus de ARN dado a que sus proteínas están emparentadas con algunos virus de ARN.[31][9] Los virus de ARN (Orthornavirae) y Duplodnaviria no formaron clados con ningún elemento egoísta, esto sugiere que son descendientes directos del mundo de ARN. Los virus de ARN serían los primeros replicadores de ARN con proteínas que aparecieron en el mundo de ARN junto con los ribosomas.[35][36]
En 2019 se propuso una hipótesis llamada "quimérica" para el origen de los virus. Esta hipótesis sostiene que los virus se originaron de distintos tipos de replicadores primordiales (protovirus) del mundo de ARN. Los virus retienen un módulo de replicación al parecer heredado de la etapa prebiótica ya que este esta ausente en las células. Según esta hipótesis al momento en que aparecieron los protobiontes estos replicadores dieron origen a los virus (viriones) al robar proteínas de sus huéspedes pre-celulares para la formación de cápsides y a su vez los virus también pudieron continuar robando proteínas de sus huéspedes celulares actuales. Estas proteínas robadas son las que conforman las enzimas virales como las polimerasas que son las que muestran más similitudes con los organismos celulares que el totalidad de proteínas expresadas (proteomas). También las infecciones virales produjeron el desplazamiento de genes asociados con la formación de cápsides y el módulo de replicación vírico entre los organismos celulares que pudieron fomentar a los elementos genéticos móviles (plásmidos, transposones de ADN, retrotransposones y los intrones del grupo II) a originar determinados virus a lo largo de la evolución.[37]
Los virus y agentes subvirales desde su aparición pudieron haber hecho contribuciones genómicas y estructurales a los organismos celulares según algunos autores. Inclusive pudieron haber originado los elementos genómicos que componen las células y haber contribuido a su formación. Por ejemplo se ha propuesto que los viroides junto con los ribozimas fueron los primeros replicadores de la etapa inicial del mundo de ARN y al haberse unido con proteínas formarían los virus de ARN, satélites de ARN junto con los ribosomas, posteriormente los virus de ARN y satélites de ARN originarian los retroelementos como los intrones del grupo II y otros, a su vez estos elementos harían la transición del ARN al ADN dando origen a los virus de ADN, satélites de ADN, transposones y plásmidos. Estos acelulares o elementos genéticos al haberse unido con liposomas formados espontáneamente originarían los protobiontes menos complejos y estos por último originarían al último antepasado común universal.[38][36][39][40][41] Sin embargo para los autores lo más probable es que los virus y satélites de la etapa prebiótica no hayan sido viriones (virus con cápsides y envoltura vírica) sino que más bien serían cadenas de ARN o ADN autorreplicantes "víricas". La aparición de los viriones coincidiría con la de los protobiontes.[37][38] Otra teoría que se ha propuesto es que los virus gigantes pudieron haber originado el núcleo de las células eucariotas al haberse incorporado el virus dentro de la célula donde en lugar de replicarse y destruir la célula huésped, permanecería dentro de la célula originando posteriormente el núcleo y dando lugar a otras innovaciones genómicas. Esta teoría es conocida como la "eucariogénesis viral".[42]
La filogenia entre órdenes virales serían las siguientes:
| Riboviria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Estudios de los pliegues de las proteínas virales y los proteomas de varias familias virales y varios filos celulares mostraron que los acelulares son un taxón complejamente parafilético.[43] El análisis demostró que los virus de ARN formaron un grupo parafilético de los virus de ADN como bien se había sugerido anteriormente, pero los organismos celulares (Cytota) surgieron dentro los virus de ADN siendo la familia Mimiviridae la más cercana a estos. Los virus retrotranscritos y los virus gigantes no formaron grupos monofiléticos lo que implica que evolucionaron independientemente en diferentes líneas evolutivas. Los autores proponen que lo más probable es que los virus se hallan originado en protobiontes que se separaron para dar origen a los procariotas o también es probable que se hayan originado de replicadores primordiales durante el mundo de ARN.[44]
La filogenia fue la siguiente:[9][10][45]
| Acytota |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Véase también
- Virus (para información sobre virus)
- Protobionte
- Sistema de dos imperios
Referencias
- ↑ a b (en inglés) Steenhuysen, Julie (20 de mayo de 2009). «Study turns back clock on origins of life on Earth». Reuters. Consultado el 27 de septiembre de 2014.
- ↑ «Virus Taxonomy: 2019 Release» (html). International Committee on Taxonomy of Viruses (ICTV) (en inglés). October 2018. Consultado el 13 octobre 2019.
- ↑ Kango, Naveen (2013). Textbook of Microbiology. I. K. International. p. 128. ISBN 9380026447.
- ↑ Dupre, John (2012). Processes of Life: Essays in the Philosophy of Biology. OUP Oxford, 2012. p. 214. ISBN 0199691983.
- ↑ «Virus Taxonomy: 2019 Release» (html). International Committee on Taxonomy of Viruses (ICTV) (en inglés). October 2018. Consultado el 13 octobre 2019.
- ↑ a b Error en la cita: Etiqueta
<ref>no válida; no se ha definido el contenido de las referencias llamadasncbi - ↑ Institut de Génétique et Microbiologie (abril de 2006). «The origin of viruses and their possible roles in major evolutionary transitions.». Virus Res. 5 (117): 5-16. PMID 16476498. Consultado el 25 de junio de 2010.
- ↑ The distribution and impact of viral lineages in domains of life. Frontiers.
- ↑ a b c d e Arshan, Nasir; Caetano-Anollés, Gustavo (25 de septiembre de 2015). «A phylogenomic data-driven exploration of viral origins and evolution». Science Advances 1 (8): e1500527. Bibcode:2015SciA....1E0527N. PMC 4643759. PMID 26601271. doi:10.1126/sciadv.1500527.
- ↑ a b c Yates, Diana (25 de septiembre de 2015). «Study adds to evidence that viruses are alive». Champaign, IL: University of Illinois at Urbana–Champaign. Archivado desde el original el 19 de noviembre de 2015. Consultado el 20 de octubre de 2015. Parámetro desconocido
|url-status=ignorado (ayuda); Parámetro desconocido|df=ignorado (ayuda) - ↑ Viroids and Virusoids Relics of RNA World Scielo.
- ↑ Viroids: “living fossils” of primordial RNAs? NCBI.
- ↑ a b Koonin, E.V.; Senkevich, T.G.; Dolja, V.V. (2006). «The ancient Virus World and evolution of cells». Biology Direct 1: 29. PMC 1594570. PMID 16984643. doi:10.1186/1745-6150-1-29.
- ↑ a b Viruses and cells intertwined since the dawn of evolution - NCBI
- ↑ Agents subvirals Sciencedirect
- ↑ 2018 ICTV Newsletter #15, September 2018 International Committee on Taxonomy of Viruses. Consultado el 26 de septiembre de 2018.
- ↑ Dávila-Ramos S, Castelán-Sánchez HG, Martínez-Ávila L, Sánchez-Carbente MD, Peralta R, Hernández-Mendoza A, Dobson AD, Gonzalez RA, Pastor N, Batista-García RA (2019). «A Review on Viral Metagenomics in Extreme Environments». Frontiers in Microbiology 10: 2403. PMC 6842933. PMID 31749771. doi:10.3389/fmicb.2019.02403.
- ↑ Baltimore, D. (1974). «The strategy of RNA viruses». Harvey Lect. 70 Series: 57-74. PMID 4377923.
- ↑ Clasificación de Baltimore
- ↑ David R. Wessner (2010). The Origins of Viruses. Nature Education.
- ↑ Institut de Génétique et Microbiologie (abril de 2006). «The origin of viruses and their possible roles in major evolutionary transitions.». Virus Res. 5 (117): 5-16. PMID 16476498. Consultado el 25 de junio de 2010.
- ↑ McClintock, B. (junio de 1950). «The origin and behavior of mutable loci in maize». Proc Natl Acad Sci U S A. 36 (6): 344-55. PMC 1063197. PMID 15430309. doi:10.1073/pnas.36.6.344.
- ↑ Do Viruses Exchange Genes across Superkingdoms of Life?. Frontiers.
- ↑ Edward N Trifonov & Eduard Kejnovsky 2015, Acytota – associated kingdom of neglected life Journal of biomolecular Structure & Dynamics 34(8):1-13 DOI: 10.1080/07391102.2015.1086959
- ↑ Viral evolution: Primordial cellular origins and late adaptation to parasitism. NCBI.
- ↑ Feschotte, Cédric; Gilbert, Clement (March 2012). «Endogenous viruses: insights into viral evolution and impact on host biology.». Nat Rev Genet. 13 (4): 83-96. doi:10.1038/nrg3199.
- ↑ Taylor, D. J.; J. Bruenn (2009). «The evolution of novel fungal genes from non-retroviral RNA viruses». BMC Biology 7. doi:10.1186/1741-7007-7-88.
- ↑ https://www.elconfidencial.com/tecnologia/2014-10-19/virus_264212/
- ↑ Krupovic, Mart; Bamford, Dennis H; Koonin, Eugene V (2014). «Conservation of major and minor jelly-roll capsid proteins in Polinton (Maverick) transposons suggests that they are bona fide viruses». Biology Direct 9 (1): 6. PMC 4028283. PMID 24773695. doi:10.1186/1745-6150-9-6.
- ↑ Natalya Yutin, Eugene Koonin, Didier Raoult (2014). Virophages, Polintons, and Transpovirons: A Complex Evolutionary Network of Diverse Selfish Genetic Elements With Different Reproduction Strategies. Virology Journal.
- ↑ a b Koonin EV, Dolja VV, Krupovic M, Varsani A, Wolf YI, Yutin N, Zerbini M, Kuhn JH (18 October 2019). «Create a megataxonomic framework, filling all principal taxonomic ranks, for realm Riboviria» (docx). International Committee on Taxonomy of Viruses (ICTV) (en inglés). Consultado el 15 de mayo de 2020.
- ↑ Koonin EV, Dolja VV, Krupovic M, Varsani A, Wolf YI, Yutin N, Zerbini M, Kuhn JH (18 October 2019). «Create a megataxonomic framework, filling all principal/primary taxonomic ranks, for dsDNA viruses encoding HK97-type major capsid proteins» (docx). International Committee on Taxonomy of Viruses (en inglés). Consultado el 19 de mayo de 2020.
- ↑ Eugene Koonin, Valerian V Doljja (2014). A virocentric perspective on the evolution of life. Science Direct.
- ↑ a b Eugene V Koonin, Tatiana G Senkevich, Valerian V Dolja (2006). The ancient Virus World and evolution of cells. Biology Direct.
- ↑ a b Krupovic, M; Dolja, VV; Koonin, EV (2019). «Origin of viruses: primordial replicators recruiting capsids from hosts.». Nature Reviews Microbiology 17 (7): 449-458. PMID 31142823. doi:10.1038/s41579-019-0205-6.
- ↑ a b Eugene Koonin, Valerian V Doljja (2014). Virus World as an Evolutionary Network of Viruses and Capsidless Selfish Elements. Microbiology and Molecular Biology Reviews.
- ↑ Eugene Koonin (2015). Viruses and mobile elements as drivers of evolutionary transitions. NCBI.
- ↑ Patrick Forterre. The Two Ages of the RNA World, and the Transition to the DNA World: A Story of Viruses and Cells. Science Direct.
- ↑ Did DNA Come From Viruses?.
- ↑ Philip John Livingstone Bell (2001). «Viral eukaryogenesis: Was the ancestor of the nucleus a complex DNA virus?». Journal of Molecular Evolution 53 (3): 251-256. Bibcode:2001JMolE..53..251L. PMID 11523012. doi:10.1007/s002390010215.
- ↑ Rijja Hussain Bokhari, Nooreen Amirjan, Hyeonsoo Jeong, Kyung Mo Kim, Gustavo Caetano-Anollés, Arshan Nasir (2020). Bacterial Origin and Reductive Evolution of the CPR Group. Oxford Academic. https://doi.org/10.1093/gbe/evaa024
- ↑ Nasir, Arshan; Kim, Kyung Mo; Caetano-Anollés, Gustavo (1 de septiembre de 2012). «Viral evolution». Mobile Genetic Elements 2 (5): 247-252. ISSN 2159-2543. PMC 3575434. PMID 23550145. doi:10.4161/mge.22797.
- ↑ Giant viruses coexisted with the cellular ancestors and represent a distinct supergroup along with superkingdoms Archaea, Bacteria and Eukarya BMC
{kind=link}
| Control de autoridades |
|
|---|
 Datos: Q169731
Datos: Q169731 Multimedia: Non-cellular life / Q169731
Multimedia: Non-cellular life / Q169731 Especies: Acytota
Especies: Acytota