
Terminología descriptiva de las flores y de los frutos (Botánica)
- Artículo introductorio en Terminología descriptiva de las plantas#Introducción a los órganos de la planta
Los términos de la morfología floral ocupan un lugar significativo en la descripción de los taxones de plantas. Ya fueron introducidos en el artículo Terminología descriptiva de las plantas. Este artículo ampliará con definiciones de términos que son útiles como atributos para diferenciar taxones.
Sexo de la flor y sexo de la planta[editar]
Por sexo de la flor nos referimos a la presencia o ausencia de partes florales masculinas o femeninas dentro de la flor. Muchas flores son perfectas (bisexuales, monoclinas, hermafroditas), tienen estambres y carpelos. Es probablemente la condición ancestral en angiospermas.
Muchos taxones de angiospermas tienen flores imperfectas (unisexuales, diclinas). En este caso, las flores pueden ser femeninas (solo carpelos) o masculinas (solo estambres). También se las puede llamar flores carpeladas o pistiladas, y flores estaminadas. Todas las flores unisexuales son además incompletas (les falta algún verticilo floral), pero puede haber flores incompletas y bisexuales (como las que no tienen perianto).
Existen algunos casos en que la flor no presenta ninguno de los verticilos reproductivos, y solo muestra sépalos y pétalos. La función de estas flores es el de especializarse, dentro de un grupo de flores que sí poseen verticilos reproductivos, en la atracción de los insectos polinizadores hacia la inflorescencia. Tales flores se denominan neutras o asexuales, se disponen usualmente en la periferia de la inflorescencia y se pueden observar, por ejemplo, en muchas especies de la familia de las compuestas, como la margarita (Bellis perennis) o el girasol (Helianthus annuus).[1]
-
 Flor hermafrodita.
Flor hermafrodita. -
 Flor masculina.
Flor masculina. -
 Flor femenina.
Flor femenina. -
 Flor asexual.
Flor asexual.




Sexo de la planta hace referencia a la presencia y distribución de las flores perfectas o imperfectas en los individuos de una especie. Una planta hermafrodita tiene solo flores bisexuales. Una planta monoica tiene solo flores unisexuales, pero tiene flores tanto masculinas como femeninas en el mismo individuo, por ejemplo en Quercus. Una planta dioica tiene solo flores unisexuales, pero cada individuo tiene solo flores masculinas o solo flores femeninas, por ejemplo en Salix. El sexo de la planta puede variar entre individuos de la misma especie, y a veces puede haber una combinación de flores bisexuales y unisexuales en el mismo individuo. Una planta polígama es una planta que tiene flores unisexuales y bisexuales. Una planta andromonoica es la que tiene flores perfectas y masculinas en el mismo individuo. Una planta ginomonoica es la que tiene flores perfectas y femeninas en el mismo individuo. Una planta trimonoica es la que tiene flores perfectas, masculinas y femeninas en el mismo individuo. Una planta androdioica es la que tiene individuos con flores masculinas e individuos con flores perfectas. Una planta ginodioica es la que tiene individuos con flores femeninas e individuos con flores perfectas. Una planta trioica es la que tiene individuos con flores femeninas, individuos con flores masculinas, e individuos con flores perfectas. Todos estos mecanismos de sexo de la planta no hermafrodita pueden ser un mecanismo para promover la fecundación cruzada entre individuos de la especie.
El control de la expresión del sexo es genético, pero puede haber un factor ambiental.
Inserción de la flor al tallo[editar]
La flor puede ser pedicelada, si tiene pedicelo, o sésil, si no lo tiene, o subsésil, si tiene un pedicelo corto y rudimentario. La flor es bracteada si está en la axila de una bráctea y no bracteada si no lo está.
-
Flor pedicelada, bracteada.
-
 Flor sésil, bracteada.
Flor sésil, bracteada.

Verticilos de la flor[editar]
Los dos términos son flor completa, si tiene todos sus verticilos florales (sépalos, pétalos, estambres y carpelos), o incompleta, si le falta alguno o más de estos verticilos (por ejemplo todas las flores unisexuales son incompletas, como también las que no tienen perianto).
Simetría floral[editar]
Se introdujo la simetría floral en #Flor.
La simetría floral se refiere a la presencia y número de planos de simetría en una flor en vista distal. La simetría radial (flor radiada, actinomórfica, regular, polisimétrica) es aquella en la que hay 3 o más planos de simetría, de forma que hay una estructura morfológica que se repite en cada porción delimitada por dos radios. La simetría birradial tiene dos y solo dos planos de simetría perpendiculares (a veces la diferencia no es reconocida, llamando a las dos simetría radial). La simetría bilateral (flor zigomórfica, monosimétrica) es aquella que tiene un único plano de simetría, entonces la flor tiene dos imágenes especulares.
Todos los casos mencionados hasta ahora, en que la flor tiene algún plano de simetría, poseen flores cíclicas.[2] Una flor asimétrica o acíclica es la que no tiene plano de simetría, usualmente como resultado de la torcedura de algunas de sus partes, como Maranta.
Maduración de la flor y la inflorescencia[editar]
Por maduración de la flor entendemos al momento en que la flor se vuelve funcional. Se llama antesis al momento de la apertura de las flores, que deja expuestas sus partes para la polinización. Se denomina protandria si la apertura de las anteras (la dehiscencia de las anteras), y la consecuente liberación del polen, ocurre antes de que los estigmas se vuelvan receptivos (hagan germinar el polen recién depositado). Se denomina protoginia a lo contrario. Los dos pueden funcionar para promover la fecundación cruzada entre individuos.
"Maduración centrífuga" y "maduración centrípeta" en el artículo Terminología descriptiva de las plantas.
Perianto[editar]
Arreglo del perianto, ciclos, merosidad[editar]
Se vio "verticilado" y "espiralado" en Terminología descriptiva de las plantas.
Normalmente un perianto, como ancestralmente, es biseriado, diclamídeo. Esto quiere decir que tiene dos verticilos, y que son distintos (cáliz y corola). Por ciclos se hace referencia al número de verticilos del perianto. El tipo más común es el perianto biseriado o dicíclico, en el que hay dos verticilos discretos, uno externo y uno interno. Una forma menos común es el perianto uniseriado con las partes del perianto en un solo verticilo. Los periantos uniseriados pueden aparecer por la pérdida de uno de los verticilos del perianto. Si se sabe que lo que se perdió fueron los sépalos, lo que quedó fue la corola, si se perdieron los pétalos, lo que quedó es el cáliz. Si no se sabe cuál verticilo se perdió, normalmente el verticilo que queda se llama cáliz por tradición. Raramente el perianto puede ser triseriado o tricíclico (con tres verticilos), o tetraseriado o tetracíclico (con cuatro verticilos). El término multiseriado puede ser usado para referirse a un perianto con 3 o más verticilos. Aclamídeo o aperiantado o flor desnuda significa que no tiene perianto.
Términos que evalúan la similitud de los verticilos entre sí. Un perianto diclamídeo tiene cáliz externo y corola interna, en la mayoría de los casos el perianto diclamídeo también es biseriado, pero podría ser multiseriado. Un perianto homoclamídeo o monoclamídeo es el que tiene todos los verticilos iguales, cada parte es un tépalo. La mayoría de las monocotiledóneas tiene un perianto homoclamídeo. En algunos casos la distinción entre homoclamídeo y diclamídeo puede ser dificultosa. Apétalo es que no tiene pétalos, asépalo que no tiene sépalos.
-
Flor periantada. Perianto biseriado, diclamídeo.
-
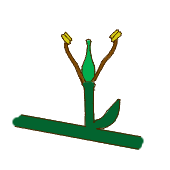 Flor aperiantada (sin perianto).
Flor aperiantada (sin perianto). -
 Flor apétala.
Flor apétala. -
 Flor asépala.
Flor asépala.



Normalmente un perianto es isómero, tiene el mismo número de piezas en cada verticilo. Por merosidad entendemos al número de piezas por verticilo. La merosidad del perianto es el número de piezas en cada verticilo del perianto. Términos generales incluyen al perianto isómero, en que los diferentes verticilos tienen el mismo número de piezas, y anisómero, en que tienen diferente número de piezas. También puede hablarse de la merosidad del cáliz y la merosidad de la corola. Si hay fusión de piezas, la merosidad se refiere al número de lóbulos. Otros términos son bímero (con dos piezas), trímero (con tres piezas), tetrámero (con cuatro piezas), pentámero (con 5 piezas).
Fusión del perianto[editar]
Ancestralmente el perianto es dialisépalo y dialipétalo, tiene los sépalos y pétalos sin fusionar entre ellos en cada verticilo. Este carácter puede ser tratado separadamente como fusión del cáliz o fusión de la corola. Si los sépalos, los pétalos o los tépalos son discretos y sin fusionar, los términos respectivos son dialisépalo, dialipétalo y dialitépalo ("aposepalous", "apopetalous" y "apotepalous" en inglés). Si parecen estar fusionados entre sí (conados), aunque sea solo en la base, los términos respectivos son gamosépalo, gamopétalo y gamotépalo ("sinsepalous", "sinpetalous" y "sintepalous" en inglés). La "fusión" de las partes del perianto no ocurre después de que las piezas fueron formadas, típicamente es el resultado del crecimiento de un primordio floral común en la base del perianto. La fusión de todas las piezas de uno o más verticilos del perianto resulta en el desarrollo de una estructura como un tubo o una copa, si la fusión no ocurre todo a lo largo de la pieza, la región como un tubo solo se encuentra en la base de las piezas, y pueden encontrarse lóbulos de perianto en la parte superior que en general marcan la posición de las piezas.
-
 Perianto dialisépalo y dialipétalo(la condición ancestral).
Perianto dialisépalo y dialipétalo(la condición ancestral). -
 Perianto dialisépalo y gamopétalo.
Perianto dialisépalo y gamopétalo. -
 Perianto gamosépalo y dialipétalo.
Perianto gamosépalo y dialipétalo. -
 Perianto gamosépalo y gamopétalo.
Perianto gamosépalo y gamopétalo.




Prefloración del perianto[editar]
La prefloración o vernación del perianto ("aestivation" en inglés) está definida por la posición, el arreglo, y la superposición de las piezas florales del perianto. La prefloración puede ser un carácter sistemático importante para delimitar o diagnosticar algunos grupos de angiospermas. La prefloración se observa mejor haciendo secciones de yemas florales maduras (para lo que se necesita un bisturí de hoja delgada y si la flor es pequeña, una lupa de disección) porque después de la antesis, la prefloración del perianto puede quedar oscurecida. Para flores muy pequeñas, la sección histológica puede ser necesaria para verla claramente.
Algunos términos estandarizados son los siguientes:
- Imbricada, un término general para las piezas del perianto que se superponen.
- Imbricada-alternada, imbricada con el verticilo externo del perianto alternando con las piezas internas del perianto.
- Convoluta o contorta, imbricada con las piezas de un único verticilo que se superponen en un margen, siendo superpuestas en el otro, como en las corolas de muchas Malvaceae.
- Quincuncial, imbricada con las piezas del perianto de un único verticilo pentámero teniendo dos piezas que superponen a otras en los dos márgenes, y una superponiendo a otra en un margen.
- Valvar, con un verticilo de piezas del perianto encontrándose en los márgenes sin superposición.
- Involuta, valvado con cada pieza del perianto induplicada (doblada longitudinalmente hacia adentro).
- Ver prefloración vexilar en Terminología descriptiva de las plantas#Glosario de Fabaceae
-
 Prefloración imbricada-alternada.
Prefloración imbricada-alternada. -
 Prefloración convoluta o contorta.
Prefloración convoluta o contorta. -
 Prefloración quincuncial.
Prefloración quincuncial. -
 Prefloración valvar.
Prefloración valvar. -
 Prefloración involuta (probablemente un error de Simpson (2005, 2010), esta postura longitudinal es reduplicada).
Prefloración involuta (probablemente un error de Simpson (2005, 2010), esta postura longitudinal es reduplicada).





Estambres[editar]
Arreglo de los estambres, ciclos, número y posición[editar]
El arreglo de los estambres es principalmente la posición que ocupan uno respecto al otro. Dos tipos básicos son:
- la disposición espiralada (en espiral), con los estambres dispuestos en espiral,
- y la disposición verticilada, con los estambres en uno o más verticilos (en general solo uno o dos).
Adicionales tipos de arreglos consideran la longitud relativa de los estambres:
- dídimos, con los estambres en dos pares del mismo tamaño,
- didínamos, con los estambres en dos pares de diferente tamaño, como en muchas Bignoniaceae, Lamiaceae, Scrophulariaceae, etc.
- tetradínamos, con los estambres en tres pares: dos grupos de dos estambres largos y un grupo de dos estambres cortos, el arreglo típico de Brassicaceae.
-
 Estambres dídimos.
Estambres dídimos. -
 Estambres didínamos.
Estambres didínamos. -
 Bignoniácea, estambres en un arreglo didínamo.
Bignoniácea, estambres en un arreglo didínamo. -
 Estambres tetradínamos.
Estambres tetradínamos.




Los ciclos de los estambres son el número de verticilos presentes (se aplica solo a los estambres de disposición verticilada). Los dos tipos principales son
- uniseriados, con un único verticilo de estambres, o
- biseriados, con dos verticilos de estambres.
- Si hay verticilos adicionales se los puede llamar triseriados, tetraseriados, etc.
Lo más común es que los estambres sean biseriados (en dos verticilos), diplostémonos (el verticilo externo antesépalo y el interno antepétalo).
- La posición de los estambres es su ubicación con respecto al radio en que se ubican otras piezas florales no estambres. En los estambres uniseriados, una posición antisépala u opositisépala es una en la cual los estambres están opuestos a los sépalos (en línea con ellos) o con los lóbulos del cáliz. Una posición alternipétala es una en la cual los estambres se alternan con los pétalos (no están en línea con ellos) o con los lóbulos de la corola. Usualmente estos dos términos son sinónimos porque usualmente el cáliz está alternado con la corola. Esta posición de los estambres es la más usual en las flores que tienen un solo verticilo de estambres.
- Un verticilo de estambres puede posicionarse en relación con el radio donde se ubican sépalos y pétalos:
- antesépalos, antisépalos u opositisépalos están ante los sépalos, opuestos a ellos, en su mismo radio, o con los lóbulos que marcan su posición.
- antepétalos, antipétalos u opositipétalos si ante los pétalos, opuestos a los pétalos (en el mismo radio, en línea con ellos) o con los lóbulos de la corola.
- alternisépalos si se posicionan alternados a los sépalos (no en línea con ellos),
- alternipétalos si se posicionan alternados a los pétalos (no en línea con ellos).
-
 La posición más usual en flores con un solo verticilo de estambres: Antesépalos, alternipétalos. Es el arreglo que queda cuando el grupo pierde el verticilo interno de estambres.
La posición más usual en flores con un solo verticilo de estambres: Antesépalos, alternipétalos. Es el arreglo que queda cuando el grupo pierde el verticilo interno de estambres. -
 Estambres uniseriados antepétalos, alternisépalos.
Estambres uniseriados antepétalos, alternisépalos.


- Dos tipos de posiciones se aplican a los estambres biseriados (en dos verticilos):
- diplostémona, en que el verticilo externo es opuesto a los sépalos y el interno es opuesto a los pétalos, y
- obdiplostémona, en el caso contrario.
- La posición diplostémona es la más común, la posición obdiplostémona es relativamente rara, siendo diagnóstica de por ejemplo, algunas Crassulaceae.
-
 Estambres biseriados diplostémonos. La posición más común en flores con dos verticilos de estambres. "External": verticilo externo.
Estambres biseriados diplostémonos. La posición más común en flores con dos verticilos de estambres. "External": verticilo externo. -
 Estambres biseriados obdiplostémonos. "External": verticilo externo.
Estambres biseriados obdiplostémonos. "External": verticilo externo.


- monandras si tienen un solo estambre, como algunas euforbiáceas
- diandras si tienen dos estambres, como las oleáceas
- poliandras si numerosos estambres, como las mirtáceas.
- El número de estambres puede ser igual o no al número de pétalos. Así, se dice que la flor es:
- isostémona si presenta el mismo número de estambres que de pétalos
- anisostémona si es diferente la cantidad de pétalos que de estambres (por ejemplo, en el género Brassica, hay cuatro pétalos y 6 estambres);
- diplostémona (ya fue definido en estambres en dos verticilos)
- polistémona cuando el número de estambres es más que el doble de pétalos
- Estos términos no son sinónimos de los anteriores, no mencionan su ubicación en verticilos.
Gineceo[editar]
Definido en Terminología descriptiva de las plantas.
- Fusión del gineceo
El grado de fusión de los carpelos, cuando son más de uno, es un carácter importante taxonómicamente y define grupos en las categorías de género y familia.
- Un gineceo apocárpico o dialicarpelar es el que tiene los carpelos separados, cada uno formando un pistilo (como en la rosa, Rosa, en Alisma, o en la magnolia, Magnolia). El término no se aplica en flores con un solo carpelo. Un gineceo dialicarpelar es probablemente la condición ancestral en angiospermas.
- Un gineceo sincárpico o gamocarpelar es el que tiene carpelos fusionados en un pistilo compuesto, y es el tipo más común en angiospermas. Los carpelos pueden estar conados solo en el ovario y tener estilos y estigmas libres (una condición muy común), o pueden estar conados solo en la base y entonces el ovario estar fuertemente lobado, o pueden estar conados en toda su extensión en un único ovario, estilo y estigma. La fusión de los carpelos determina el tipo de placentación.
- Número de carpelos y lóculos
El número de carpelos y lóculos es importante taxonómicamente. El número de lóculos es en general fácil de determinar en una sección transversal o longitudinal cuidadosa del gineceo, ya que es equivalente al número de cámaras cerradas dentro del ovario. En general, los ovarios pueden ser uniloculares, con un único lóculo, o pluriloculares, con más de un lóculo. En algunas angiospermas, los septos pueden dividir al ovario en cámaras en una región, como la base del ovario, pero no en otra región, como la parte apical del ovario. En ese caso, las cámaras de abajo son continuas con la única cámara de arriba, y el ovario es unilocular.
El número de carpelos es muchas veces crítico en la clasificación y la identificación de angiospermas. Se determina como sigue. Si el gineceo es dialicarpelar, el número de carpelos es equivalente al número de pistilos. Si hay un único pistilo, puede estar formado por uno o varios carpelos. Para un único pistilo el número de carpelos se determina como sigue, en secuencia: Primero, el número de carpelos es igual al número de estilos o estigmas, si alguno de éstos está en un número mayor a 1. Esto es así más allá de la estructura del carpelo porque los estilos o estigmas son interpretados como estructuras de carpelos vestigiales. Por ejemplo, los miembros de Asteraceae tienen dos estilos y dos estigmas, por lo tanto se interpreta que las asteráceas tienen dos carpelos. Esto es así aunque haya un solo lóculo, óvulo, y placenta, que se interpreta que son únicos por reducción en la evolución. Segundo, si un pistilo tiene un solo estilo y un solo estigma, el ovario tiene que ser disectado para revelar el número de carpelos. Si el ovario es plurilocular, entonces el número de lóculos generalmente se corresponde con el número de carpelos. Cada lóculo representa la cámara de un carpelo, excepto en algunos taxones. Finalmente si el ovario es unilocular, el número de carpelos es igual al número de placentas. Por ejemplo una violeta, con un pistilo, un estilo, un estigma, y un lóculo, se interpreta que tiene 3 carpelos porque tiene 3 placentas (con placentación parietal). Excepciones a las dos últimas reglas son los taxones ginobásicos de Lamiaceae y Boraginaceae sensu stricto, en estos dos grupos cada uno de los dos carpelos es biseccionado temprano en el desarrollo por un llamado falso septo, de forma que el ovario maduro tiene típicamente 4 lóculos, cada uno con una placenta y un óvulo. En este caso no se aplican las reglas descriptas.
En Malus sylvestris, el manzano, la soldadura de los carpelos es incompleta, de modo que se forma un único lóculo estrellado, visible en el corte transversal de su fruto, la manzana. Excepcionalmente los carpelos pueden disponerse en 2-3 ciclos o pisos como sucede en Punica granatum (la granada).[5]
- Posición con respecto al perianto y androceo
Si bien los términos que siguen son muy usados, pueden ser reemplazados por la posición del ovario y la presencia o ausencia de hipanto (ovario súpero o ínfero, o semiínfero, con o sin hipanto), en Terminología descriptiva de las plantas.
Tres términos describen a la flor sin hipanto: el ovario hipógino es usado para aquel que tiene a los sépalos, pétalos y estambres unidos a la base de un ovario que por lo tanto es súpero. Un ovario epígino tiene los sépalos, pétalos y estambres unidos al ápice de un ovario que por lo tanto es ínfero. El término epihipógino se usa para el que los tiene unidos al medio del ovario.
Otros términos describen al ovario de la flor con hipanto, con los sépalos, pétalos y estambres unidos al hipanto. Un ovario perígino denota al ovario con el hipanto unido a la base de un ovario que por lo tanto es súpero. Un ovario epiperígino tiene al hipanto unido al ápice de un ovario que por lo tanto es ínfero. El extraño término epihipoperígino puede usarse para el ovario con el hipanto unido en el medio de un ovario que por lo tanto es semiínfero.
- Placentación
La placentación es la ubicación de los óvulos y tiene en cuenta el número de placentas y su ubicación, de los septos, y de los lóculos. Determinar la placentación requiere observar el ovario a fondo haciendo un corte longitudinal o transversal de él.
Tipos de placentación estándar son la placentación axilar, en que la placenta se encuentra en una columna en un ovario compuesto con septos, común en muchas angiospermas como Liliaceae, la placentación apical o péndula, con la placenta en el ápice del ovario, la placentación apical-axilar, con dos o más placentas en la punta de un ovario septado, como en Apiaceae, la placentación basal, con una placenta en la base del ovario, como en Asteraceae y Poaceae, la placentación libre-central, con la placenta a lo largo de una columna en un ovario compuesto sin septos, como en Caryophyllaceae, la placentación laminar o parietal difusa, con óvulos saliendo de la superficie de los septos, la placentación marginal, con las placentas a lo largo del margen de un ovario unicarpelar, como en Fabaceae, la placentación parietal, con las placentas en las paredes del ovario o en las particiones que se introducen en un ovario compuesto unilocular, como en Violaceae, la placentación parietal-axilar, con las placentas en la unión entre los septos y la pared del ovario en un ovario plurilocular, como en Brassicaceae, y la placentación parietal-septada, con las placentas en las paredes internas del ovario entre los septos de los lóculos, como en algunas Aizoaceae.
La placentación axilar puede contener placentas intrusivas (un crecimiento de la placenta que interna a los óvulos aún más dentro del lóculo), lo que se llama placentación axilar con placentas intrusivas o "placentación axilar con doble curvatura de los carpelos".[5]
-
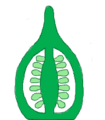 Placentación axilar, corte longitudinal.
Placentación axilar, corte longitudinal. -
 Placentación axilar con dos lóculos, corte transversal.
Placentación axilar con dos lóculos, corte transversal. -
 Placentación axilar con 5 lóculos, corte transversal.
Placentación axilar con 5 lóculos, corte transversal. -
 Placentación apical o péndula, corte longitudinal.
Placentación apical o péndula, corte longitudinal.




-
 Placentación apical-axilar, corte longitudinal.
Placentación apical-axilar, corte longitudinal. -
 Placentación basal, corte longitudinal.
Placentación basal, corte longitudinal. -
 Placentación libre-central, corte transversal.
Placentación libre-central, corte transversal. -
 Placentación laminar, corte transversal.
Placentación laminar, corte transversal.




-
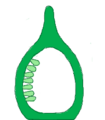 Placentación marginal, corte longitudinal.
Placentación marginal, corte longitudinal. -
 Placentación marginal, corte transversal.
Placentación marginal, corte transversal. -
 Placentación parietal, corte transversal.
Placentación parietal, corte transversal. -
 Placentación parietal (con placentas intrusivas), corte transversal.
Placentación parietal (con placentas intrusivas), corte transversal.




-
 Placentación parietal-axilar, corte transversal.
Placentación parietal-axilar, corte transversal. -
 Placentación parietal-septada, corte transversal.
Placentación parietal-septada, corte transversal. -
 Placentación axilar con placentas intrusivas, corte transversal.
Placentación axilar con placentas intrusivas, corte transversal.



- Estilo
Los estilos pueden estar estructuralmente especializados en algunos taxones. Una de estas especializaciones es el "beak" estilar, una región basal del estilo persistente, extendida, es típicamente acrescente y se elonga durante la formación del fruto. Funciona en la dispersión del fruto, como en miembros de Asteraceae (por ejemplo Taraxacum, el diente de león) o en Geraniaceae (por ejemplo en Geranium).
- Estigma
Según Heslop Harrison y Shivanna (1977[6]), los estigmas se dividen en 2 grandes grupos: estigmas húmedos y secos. Los estigmas húmedos tienen secreción presente durante el período receptivo, los estigmas secos no tienen secreciones líquidas, sino que producen proteínas o ceras. Los últimos a su vez se dividen en plumosos (Gramineae) y no plumosos, los no plumosos se dividen de nuevo en papilosos (Cordyline, Yucca, Aristolochia, Bombax, Nymphaea, Pelargonium, Bougainvillea, Plumbago, Acalypha) o no papilosos (Asclepias, Capparis, Myrica, Cyperus).[7]
Términos muy usados de su morfología son estigma discoide, en forma de disco, estigma globoso o en cabezuela o capitado, en forma de globo como en Citrus, estigma lineal, con estigma o tejido estigmático largo y esbelto en forma, y estigma plumoso, con extensiones que son como tricomas que lo hacen parecer plumoso, encontrado en muchos taxones polinizados por viento (como Cyperaceae, Poaceae) (Simpson 2010), lobado en Cucurbita[7]
- Óvulo
Los óvulos, primordios seminales o rudimentos seminales nacen sobre las placentas, situadas en la cara interna del carpelo. Son de tamaño reducido, de pocos milímetros, y generalmente de forma ovoide, y autores como Simpson (2005[8]) consideran su descripción como parte del campo de la anatomía. Cada óvulo consta de un cuerpo de tejido compacto, la nucela —que representa el esporangio—, rodeada de uno o dos tegumentos. Los tegumentos son tejido esporofítico que nace de la calaza o chalaza, la región del óvulo donde convergen el funículo, la nucela y los tegumentos. El funículo es el cordón que lo une a la placenta. Los tegumentos dejan un orificio en el óvulo llamado micrópilo, que será atravesado por el tubo polínico (el crecimiento haustorial del gametófito masculino). En la superficie del óvulo, en el sector donde se termina o se fusiona al funículo, queda una cicatriz al caer el último que se llama hilo cárpico o hilo.[9]
Los óvulos se clasifican de acuerdo a las relaciones espaciales entre el funículo, la chalaza y el micrópilo:[10]
- Ortótropo (en griego ὀρθός orzós "recto", también denominado átropo o recto), es el óvulo en el cual el funículo, la chalaza y el micrópilo se hallan en la misma línea. Se considera el tipo más primitivo: Urticaceae, Piperaceae, Polygonaceae.
- Anátropo (de ἀνά aná "hacia arriba"): El cuerpo del óvulo se gira 180°, de forma que queda el micrópilo próximo al funículo. El funículo se fusiona a la cara interna de los tegumentos y desde el exterior se observa un engrosamiento alargado llamado rafe, la chalaza queda del lado opuesto al funículo. Son los óvulos más frecuentes en las angiospermas.
- Campilótropo (de καμπύλος campílos "curvo") o encorvado, es un óvulo incurvado de forma que la chalaza se halla próxima al funículo. Este tipo de óvulo es frecuente en las leguminosas (fabáceas) como se observa en los porotos.
- Tipos de óvulo
-
 Óvulo ortótropo. 1: saco embrionario (gametófito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo.
Óvulo ortótropo. 1: saco embrionario (gametófito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo. -
 Óvulo anátropo. 1: saco embrionario (gametófito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo 4: rafe.
Óvulo anátropo. 1: saco embrionario (gametófito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo 4: rafe. -
 Óvulo campilótropo. 1: saco embrionario (gametófito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo.
Óvulo campilótropo. 1: saco embrionario (gametófito femenino), rodeado del nucelo (esporangio) y los 2 tegumentos 2: calaza 3: funículo.



Inflorescencia[editar]
Definidas en Terminología descriptiva de las plantas.
Las inflorescencias parciales también se llaman paraclados si provienen de una inflorescencia determinada, y co-florescencias si provienen de una inflorescencia indeterminada.[11]
- Tipos de inflorescencia
Los tipos de inflorescencia son caracteres valiosos en sistemática y usualmente son característicos de ciertos grupos, como las umbelas compuestas de Apiaceae, las cabezas de Asteraceae, y las cimas helicoides o escorpioides de Boraginaceae. Algunos tipos de inflorescencia son bastante especializadas para reproducción, como los ciatios de Euphorbioides.
El término cima (o "tirso determinado") puede ser usado como un término general para denotar una inflorescencia determinada. Un tipo de cima es el dicasio, que desarrolla dos ejes laterales y una flor terminal, formando un par o más de ejes opuestos y laterales. Un dicasio simple o címula es una cima de 3 flores, con una única flor terminal y dos flores laterales opuestas, todos los pedicelos del mismo tamaño, típicamente con brácteas debajo de las dos flores laterales, aunque pueden estar ausentes. Un dicasio compuesto es una cima de más de 3 flores, en que los dicasios originalmente simples se ramifican varias veces en sus ejes laterales en más unidades de dicasios, las ramas están típicamente en muchos planos. Finalmente, una cima compuesta es una cima ramificada, similar a un dicasio compuesto pero sin un consistente patrón de dicasio: las cimas pueden tener diferentes formas debido a sus diferencias en sus patrones de ramificación. Algunas cimas compuestas en realidad tienen el mismo patrón de ramificación que el dicasio compuesto pero con algunos ejes reducidos o faltando, dando una apariencia más congestionada.
-
 Dicasio simple o címula.
Dicasio simple o címula. -
 Dicasio compuesto.
Dicasio compuesto.


Algunas cimas producen varios ejes laterales consecutivos que parten desde el eje principal antes de terminar en la flor terminal (se dice que la ramificación es "inicialmente monopodial"), produciendo varios entrenudos antes de llegar a la flor terminal. Estas cimas se llaman cima paniculoide o panícula de Weberling o, si cada eje lateral da una única flor por reducción, cima racimoide (la panícula y el racimo son inflorescencias con el mismo aspecto pero indeterminadas, ver figuras). Si cada eje lateral da unidades dicasios de 3 flores se llama cima elongada[11]
-
 Cima paniculoide. Nótese el desarrollo determinado.
Cima paniculoide. Nótese el desarrollo determinado.

No todas las cimas tienen ramificación opuesta (dando dos ejes opuestos), también las hay con un arreglo alternado de ejes. Un monocasio es una cima que desarrolla en cada ramificación un único eje lateral, faltando el eje opuesto (la terminología para definir a los monocasios puede variar de autor a autor, ésta es la definición dada en Simpson 2005). Una cima helicoide o bóstryx es un monocasio (cima con un único eje lateral) en el que siempre se desarrolla el eje del mismo lado en secuencia, apareciendo como una inflorescencia que se espirala al menos temprano en el desarrollo (ver figura). Una cima escorpioide o cincino es un monocasio en el cual las ramas se desarrollan en ejes alternados de cada eje secuencial, típicamente resultado en una apariencia de zig-sag (geniculada). Estos dos tipos de cimas tienen ramas o ejes en más de un plano y pueden ser vistas como derivadas por reducción de ramas decusadas (dispuestas en forma de cruz) de un dicasio compuesto. Existen otros dos tipos de cimas monocasiales que se disponen en este caso en un solo plano: el drepanio es un monocasio en el cual los ejes se desarrollan en solo un lado de cada plano secuencial, como una cima helicoide pero en un plano, típicamente aparece espiralado temprano en el desarrollo, de hecho en algunas terminologías aparecen como cimas helicoides. Un ripidio o flabelo es un monocasio en el cual las ramas se desarrollan en lados alternos de cada eje secuencial, como las cimas escorpioides pero en un plano, típicamente tienen una apariencia de zig-sag (geniculada). De hecho, los ripidios son tratados como cimas escorpioides en algunas terminologías. Estas cuatro estructuras pueden intergradarse unas con otras. Por ejemplo, un intermedio entre una cima helicoide y un drepanio puede ocurrir. Por eso algunos autores prefieren definir las cimas monocasiales sin entrar en detalles.
-
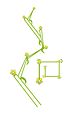 Cima helicoide o bóstryx.
Cima helicoide o bóstryx. -
 Cima escorpioide o cincino.
Cima escorpioide o cincino. -
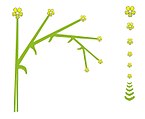 Drepanio.
Drepanio. -
 Ripidio.
Ripidio.
.jpg)

.jpg)
.jpg)
A las inflorescencias indeterminadas en general les falta la flor en el ápice del eje de la inflorescencia y se desarrollan desde la base hasta el ápice. Muchos tipos de inflorescencias indeterminadas pueden ser reconocidos. Una espiga es una inflorescencia indeterminada, en la que un único eje lleva flores sésiles. Similarmente, un racimo es una inflorescencia indeterminada en un solo eje pero lleva flores pediceladas. Una panícula es como un racimo ramificado o compuesto, definida como una inflorescencia indeterminada con muchos ejes ramificados en dos o más órdenes, que llevan flores pediceladas. Un corimbo es una inflorescencia indeterminada consistente en un eje central con ejes laterales o pedicelos (es un tipo de racimo) en el cual las flores elongan su pedicelo de forma convexa, de forma que queden todas las flores más o menos al mismo nivel y mirando hacia arriba. Los corimbos pueden ser simples o compuestos. Un corimbo simple no es ramificado, consiste en un eje central llevando flores pediceladas, la colección de flores convexas de forma que queden mirando hacia arriba, son como racimos en que los pedicelos más bajos son mucho más elongados que los superiores. Un corimbo compuesto es ramificado, consiste en dos o más órdenes de ejes de inflorescencias llevando flores convexas, pediceladas, los corimbos compuestos son como panículas en que los ejes más bajos y los pedicelos más bajos son mucho más elongados que los superiores.
-
 Corimbo simple.
Corimbo simple.

Algunas inflorescencias pueden ser determinadas o indeterminadas. Una umbela simple es determinada o indeterminada, es una inflorescencia convexa de forma que las flores queden mirando hacia arriba, en que todos los pedicelos están unidos al mismo punto del pedúnculo, por lo que todas las flores quedan al mismo nivel y con los pedicelos del mismo tamaño. Dos inflorescencias en las que las flores en el punto de unión aparecen congestionadas son el fascículo y el glomérulo. Un fascículo es una inflorescencia que puede derivar de un racimo o una panícula (o una cima[11]), con flores pediceladas, en que los entrenudos entre flores pueden ser muy cortos. Un glomérulo es una inflorescencia de flores sésiles o subsésiles en que los entrenudos entre flores son muy cortos.
-
 Fascículo.
Fascículo. -
 Glomérulo.
Glomérulo.


En algunos taxones una inflorescencia puede parecer de un tipo, pero después de un examen detallado, es en realidad una modificación de otro tipo. Por ejemplo, se puede usar el término pseudoumbela para una inflorescencia que parece una umbela simple, pero en realidad está compuesta por cimas monocasiales condensadas, como en Alliaceae y Amaryllidaceae.
- Inflorescencias secundarias
Cuando tres o más inflorescencias parciales están dispuestas en un verticilo debajo de la flor terminal, la inflorescencia se denomina pleiocasio. Ejemplos son Sedum, Sempervivum, Cornus.[12]
Dos tipos específicos de inflorescencias secundarias son el tirso y el verticiláster. El tirso (o tirso indeterminado) es esencialmente un racimo de cimas, en que el eje central es indeterminado pero las unidades de inflorescencia son cimas opuestas, laterales, y pediceladas, típicamente las unidades son dicasios simples, o dicasios compuestos, o cimas compuestas, ocasionalmente cimas monocasiales (al nombre genérico "cima" también se lo llama "tirso determinado", aquí se diferencia del "tirso" o "tirso indeterminado"). Nótese la diferencia con panícula, en que las ramas son indeterminadas. Un verticiláster es esencialmente una "espiga de cimas opuestas", similar al tirso en tener un eje central indeterminado pero diferente en que las cimas laterales tienen una apariencia muy congestionada. Los verticilásters se encuentran en muchos miembros de Lamiaceae, la familia de la menta. Una umbela compuesta es otra inflorescencia secundaria en que el pedúnculo lleva ejes secundarios llamados radios que se unen en un punto y la unidad de la inflorescencia son umbelas simples unidas al ápice de los radios, como en Apiaceae.
-
 Tirso.
Tirso. -
 Tirso.
Tirso. -
 Verticiláster.
Verticiláster. -
 Umbela compuesta.
Umbela compuesta.




- Inflorescencias especializadas
"Amento" y "cabeza" o "capítulo", "espádice", "espiguilla" en Terminología descriptiva de las plantas.
Algunas inflorescencias son bastante especializadas y muchas veces restringidas a ciertos grupos taxonómicos. Un ciatio es una inflorescencia que lleva flores pequeñas unisexuales y tiene por debajo un involucro (frecuentemente con glándulas petaloides), la inflorescencia pareciendo una única flor, como en Euphorbia y afines.La inflorescencia de Asteraceae en #Glosario de Asteraceae. Un sicono (hypanthodium en inglés) es una inflorescencia que lleva numerosas flores en la parte interior de un receptáculo muy convexo o involuto, como en Ficus, el higo. Una espiguilla es la inflorescencia básica de Cyperaceae y Poaceae. Las espiguillas de Cyperaceae son como una espiga pequeña, con flores sésiles reducidas en un eje (raquilla), cada flor tiene por debajo una bráctea llamada escama. La espiguilla de Poaceae en Terminología descriptiva de las plantas#Glosario de Poaceae
-
 Ciatio.
Ciatio. -
 Ciatio.
Ciatio. -
 Sicono.
Sicono.


.svg)
Frutos[editar]
Definidos en Terminología descriptiva de las plantas#Introducción a los órganos de la planta y en Terminología descriptiva de las plantas#Frutos.
- Frutos simples
Otras definiciones.
En el caso de los frutos derivados de ovario ínfero, la porción extracarpelar recibe el nombre de clamidocarpo. En un cierto número de especies, otras partes de la flor o de la inflorescencia se asocian al gineceo para producir el fruto. Cuando las partes de la flor que persisten en el fruto no son concrescentes con él (no están congénitamente unidas) se llaman induvias. Las brácteas también pueden formar parte del fruto como en la nuez del roble (Quercus robur) y del avellano (Corylus avellana).
Los frutos pueden tener una o varias semillas, excepcionalmente pueden no contenerlas. Si contienen solo una semilla se los llama frutos unispermos. Los frutos que no contienen semillas (y no es por accidente) son frutos partenocárpicos, como la banana comercial.[13]
La gran diversidad de tamaño, forma, textura, formas de abrirse, y anatomía entre frutos ha confundido durante mucho tiempo a los sistemáticos de plantas, y muchas clasificaciones de los frutos han sido propuestas. Todos los sistemas de clasificación de los frutos deben lidiar con muchas dificultades. Primero está el problema de la gran variación entre estructuras de frutos: van der Pijl (1972:17[14]) concluyó que "el fruto es muy versátil y tiene demasiados aspectos para ser dividido en categorías estrictas". En segundo lugar, complejidades adicionales vienen de la extensa evolución convergente de las estructuras frutales: frutos funcionalmente similares han aparecido independientemente en diferentes linajes de angiospermas de similares o diferentes condiciones del gineceo. En tercer lugar, otras partes de la flor y de las estructuras asociadas a ella, además del gineceo maduro, pueden formar parte del fruto maduro (las estructuras accesorias). Finalmente, los frutos provenientes de climas tropicales han sido poco considerados en muchos sistemas de clasificación tradicionales de los frutos. En este texto se utiliza un sistema artificial de términos descriptivos de los frutos, basado en la clasificación de los frutos tradicional de Gray (1877[15]). Este sistema ha sido extensamente utilizado. Está basado en la textura del pericarpio (carnoso, seco, o endurecido), el tipo de dehiscencia (si se abre y cómo, o si no se abre), la forma y tamaño del fruto, y el número de carpelos y semillas. Para más información acerca de los tipos de fruto, ver Judd (1985[16]), Weberling (1989[17]), y Spjut (1994[18]).[11]
Los frutos derivados de una sola flor, tanto simples como agregados, se llaman monotalámicos ("un tálamo", un solo receptáculo), los múltiples son politalámicos ("muchos tálamos").
Los frutos simples son clasificados según cierto número de criterios, los dos principales son si son carnosos o secos a la madurez, y si son dehiscentes (si la cáscara se rompe a la madurez a lo largo de poros definidos, o suturas, exponiendo el interior) o indehiscentes (que no se rompen a la madurez). El fruto carnoso suele ser una adaptación a la dispersión por los animales que se alimentan de su pulpa. En general solo los frutos secos son dehiscentes, aunque puede haber algún caso aislado de fruto carnoso y dehiscente. Además se los puede definir teniendo en cuenta (3) si dehiscentes, el tipo de dehiscencia (es decir, la ubicación, forma y dirección de dehiscencia), (4) número de carpelos y lóculos, incluyendo la presencia de septos, (5) número de semillas, (6) placentación, (7) la estructura del pericarpio, (8) posición del ovario.
Un aquenio es un fruto seco indehiscente. "Aquenio", "Nuez", "Sámara", "Grano" o "Cariópside" en Terminología descriptiva de las plantas#Frutos.
- En sentido estricto, es de una sola semilla, bastante pequeño, con un pericarpio más bien delgado y cercano a la semilla pero sin adherirse a ella. Ejemplos son Bidens, Carex, Ceratophyllum, Clematis, Cyperus, Ficus, Fragaria (la frutilla o fresa, el fruto son los aquenios, la parte comestible es el receptáculo), algunos Medicago, Ostrya, Petiveria, Polygonum, Ranunculus, Rhynchospora, Rosa (la rosa, el fruto son aquenios encerrados en un recipiente formado por el receptáculo carnoso), Rumex, Sagittaria, algunos Trifolium, Vernonia.[11]
- Un antocarpo es en general un aquenio, con la particularidad de que una o más partes de la flor funcionan como tejidos accesorios, como en Pontederia, en que un perianto acrescente rodea y se fusiona con el aquenio, o en Platanus.
- Cipsela es como se llama al aquenio de las compuestas (que posee tejido del cáliz adherido que funciona en la dispersión del fruto, por lo que también es un antocarpo), como en la unidad del fruto del girasol o del diente de león.
- Un grano o cariópside es un aquenio con el tegumento de la semilla adnato al pericarpio delgado, los granos son el tipo de fruto de casi todas las Poaceae (pastos y cereales, como trigo, maíz, etc.).
- Una nuez es un fruto simple, seco e indehiscente de una sola semilla que, en relación con el típico aquenio, es bastante grande, y su pericarpio grueso y muy duro, usualmente es derivado de un ovario de un lóculo. Ejemplos son la avellana (Corylus avellana), la castaña (Castanea), el roble (Quercus), Dipterocarpus, Fagus, Nelumbo (que posee las nueces sumergidas en el receptáculo), Shorea, Brasenia.[11] Las nueces y los aquenios pueden intergradarse, los términos a veces se usan indistintamente. Muchas nueces están rodeadas de un involucro, como la nuez de Quercus, la de Corylus cornuta o la de Castanea. Tipos más especializados de nuez:
- Una trima es una nuez rodeada de un involucro carnoso, que actúa como tejido accesorio (sin serlo, las "partes accesorias" provienen de la flor), y es dehiscente a la madurez, como en Carya y Juglans regia (según algunos autores la trima es un fruto carnoso,[13] la unidad de dispersión es carnosa). Una pseudodrupa es una nuez rodeada de un involucro carnoso e indehiscente, como en Juglans (la nuez, walnut), chequear el ejemplo.
- Una núcula es un fruto como una nuez pero pequeño, por ejemplo las pequeñas unidades que en conjunto conforman el fruto de Boraginaceae y Lamiaceae (ver las dos familias en mericarpios, más adelante).
- Una sámara es un fruto seco e indehiscente y alado, en general de una sola semilla (raramente dos). Ejemplos son Ailanthus, Betula, Casuarina, Fraxinus, Liriodendron, Myroxylon, Ptelea, Stigmaphyllon y el olmo (Ulmus).[11]
- Finalmente, un utrículo es un fruto seco de una sola semilla pequeño, con vejiga o inflado, los utrículos son esencialmente aquenios en que el pericarpio es significativamente más espacioso que la semilla madura, como en Atriplex. Más ejemplos[11] se encuentran en algunos Amaranthus, en Chenopodium, Lemna, y Limonium.
- Aquenios
-
 Aquenio, sección longitudinal.
Aquenio, sección longitudinal. -
 Grano o cariópside, sección longitudinal.
Grano o cariópside, sección longitudinal. -
 Nuez, sección longitudinal.
Nuez, sección longitudinal.



-
 Trima de nogal (Juglans regia), se observa la nuez y el involucro carnoso dehiscente.
Trima de nogal (Juglans regia), se observa la nuez y el involucro carnoso dehiscente. -
 La cáscara de la nuez es el pericarpio leñoso, el interior comestible es la semilla.
La cáscara de la nuez es el pericarpio leñoso, el interior comestible es la semilla. -
 Trima de Carya, otra juglandácea.
Trima de Carya, otra juglandácea.



-
 Sámara.
Sámara. -
 Sámaras (a la madurez se separan en unidades).
Sámaras (a la madurez se separan en unidades). -
 Utrículo, sección longitudinal.
Utrículo, sección longitudinal.



Otros frutos simples son secos y dehiscentes a la madurez. La mayoría de los frutos secos dehiscentes se abren por una valva, un poro, un opérculo, una línea de dehiscencia que expone su interior. Algunos tipos de frutos son explosivamente dehiscentes, se abren por fuerza por mecanismos variados y eyectan las semillas.
La forma más general de referirse al fruto seco dehiscente es la cápsula. Cuatro tipos de cápsulas pueden ser reconocidas mediante el tipo o ubicación de la dehiscencia. Las cápsulas loculicidas tienen líneas longitudinales de dehiscencia dispuestas radialmente con los lóculos (o entre las placentas, si no hay septos), es decir que la dehiscencia se produce a lo largo de la vena media de los carpelos ("dehiscencia loculicida"). El fruto puede permanecer entero o se fragmenta en valvas, correspondiendo cada una a las mitades de dos carpelos contiguos; cuando la placentación es axilar, cada valva presenta el septo en su parte media. Son cápsulas loculicidas los frutos de violáceas, turneráceas (frutos uniloculares) y amarilidáceas, iridáceas, Allium, Cupania y Gossypium hirsutum (el algodón), (frutos pluriloculares). Las cápsulas septicidas tienen líneas longitudinales de dehiscencia dispuestas radialmente en relación con los septos del ovario (o con las placentas, si no hay septos), como en Colchicum y Martynia (según algunos autores, cuando la cápsula se rompe por la línea media de las placentas se llama cápsula placenticida, como en Gentiana[13]). Los dos tipos de cápsulas dividen al fruto en valvas, una porción de la pared del pericarpio que se separa, pero no encierra a las semillas. Las valvas pueden quedar adheridas al fruto o pueden caer, según el taxón. La cápsula circunsisa (también llamada pixidio o "cápsula de dehiscencia transversal" o "de dehiscencia operculada") tiene una línea de dehiscencia transversa, típicamente formando un opérculo terminal, como en Plantago y Portulaca (la verdolaga). Una cápsula septífraga es la que produce valvas que se rompieron en rajaduras producidas en los septos, con las porciones internas y las semillas unidas a una columna axial, como en Ipomoea y Cedrela. Las cápsulas poricidas tienen dehiscencia que ocurre por poros ("dehiscencia poricida o foraminal"), como en Papaver (la amapola) y Antirrhinum majus (el conejito), estas cápsulas poseen un largo pedicelo que provoca que el viento las agite con facilidad, de forma que se escapen las semillas por los poros.[19] Otras cápsulas pueden ser definidas por la ubicación de la dehiscencia, como las cápsulas acrocidas ("dehiscencia dental") que se abren por hendiduras apicales formando "dientes", como en Silene italica (Caryophyllaceae) o en Eucaliptus, o las cápsulas basicidas, que se abren por hendiduras basales, como en Aristolochia (el "patito"). Hay otros tipos de frutos secos y dehiscentes que son estructuras tipo cápsula especializadas. Un folículo es un fruto seco y dehiscente derivado de un carpelo que se divide a lo largo de una única sutura longitudinal ("dehiscencia sutural simple o ventricida"), las semillas pueden ser ariladas o con un tegumento carnoso. Ejemplos son los frutos unidad de Magnolia, y también Akebia, Alstonia, Aquilegia, Asclepias, Caltha, Grevillea, Nerium, Paeonia, Sterculia, Zanthoxylum;[11] Macadamia. Cuando se combinan dos tipos de dehiscencia, como septicida y septífraga en las cápsulas de Rhododendron, o loculicida y septífraga como en las cápsulas de ovario ínfero de Belamcanda chinensis, se llama "dehiscencia biscida".[13]
- Frutos de tipo cápsula
-
 Se llama cápsulas a los frutos dehiscentes. Aquí, una inflorescencia de cápsulas aún cerradas en Liquidambar styraciflua. En este caso, no todos los pistilos maduran en una cápsula, por eso algunos son muy pequeños.
Se llama cápsulas a los frutos dehiscentes. Aquí, una inflorescencia de cápsulas aún cerradas en Liquidambar styraciflua. En este caso, no todos los pistilos maduran en una cápsula, por eso algunos son muy pequeños. -
 Cápsulas ya abiertas en Liquidambar styraciflua.
Cápsulas ya abiertas en Liquidambar styraciflua. -
 Cápsula loculicida: Fruto cerrado, fruto abierto y corte transversal con flechas mostrando el sitio de sutura.
Cápsula loculicida: Fruto cerrado, fruto abierto y corte transversal con flechas mostrando el sitio de sutura.



-
 Cápsula septicida: Corte transversal con flechas mostrando el sitio de sutura.
Cápsula septicida: Corte transversal con flechas mostrando el sitio de sutura. -
 Cápsula circuncisa o pixidio.
Cápsula circuncisa o pixidio. -
 Cápsula circuncisa o pixidio.
Cápsula circuncisa o pixidio.



-
 Cápsula poricida.
Cápsula poricida. -
 Cápsula poricida en la amapola, Papaver. El viento agita el largo pedicelo con lo cual las semillas escapan por los poros.
Cápsula poricida en la amapola, Papaver. El viento agita el largo pedicelo con lo cual las semillas escapan por los poros.


-
 Folículo, aspecto general y corte transversal.
Folículo, aspecto general y corte transversal. -
 Folículos de Brachychiton populneus.
Folículos de Brachychiton populneus.


Fabaceae. Ver "legumbre", "vaina, "lomento", "sutura longitudinal doble", en Terminología descriptiva de las plantas#Glosario de Fabaceae. Brassicaceae. "Silicua" y "silícula" en el Terminología descriptiva de las plantas#Glosario de Brassicaceae.
Frutos carnosos. "Baya y "drupa" ya definidos.
- Baya como en Vitis, la uva. En Lycopersicum esculentum (el tomate) las placentas de la baya proliferan llenando los lóculos con un tejido carnoso que envuelve totalmente las semillas, y luego sufre degeneración mucilaginosa. Actinidia chinensis (el kiwi) es una baya con semillas negras de placentación axilar, Carica papaya (el mamón) es una baya con semillas negras de placentación parietal. Algunos autores diferenciaron la baya, de ovario súpero, de la pseudobaya o baya falsa, de ovario ínfero, como la banana, pero esta diferenciación está hoy en día abandonada.
- Una drupa como en Prunus (los duraznos, la ciruela, la cereza, etc.) y ?Olea europaea (la aceituna).
- El término pireno también puede ser usado para un fruto carnoso con dos o más semillas, cada una encerrada por un endocarpio usualmente con textura de hueso (el término pireno también puede referirse a la semilla adherida al endocarpio duro solamente, más allá del número de semillas). Parece sinónimo de nuculanio,[13] por ejemplo en Mespilus germanica, el níspero.[13]
- Un hesperidio (el cítrico) es un fruto carnoso septado con un pericarpio externo grueso y coriáceo (el flavedo), mesocarpo corchoso (albedo), y tricomas modificados para ser carnosos (sacos de jugo, juice sacs) que se originan en las paredes internas, es el fruto de Citrus (naranja, limón, etc.).
- Un pepónide en el #Glosario de Cucurbitaceae al final de este artículo.
- Un pomo es un fruto carnoso con un endocarpio cartilaginoso derivado de un ovario ínfero, con la mayor parte del tejido carnoso derivado del hipanto adnato, como en Malus (la manzana) y Pyrus (la pera).
- El fruto de Punica granatum, la granada, es una balaústa: un fruto proveniente de un ovario ínfero, con pericarpio coriáceo, carpelos dispuestos en dos estratos, y semillas con episperma jugoso (sarcotesta).[13]
-
 Baya.
Baya. -
 Drupa.
Drupa. -
 Hesperidio.
Hesperidio.



-
 Pomo.
Pomo. -
 Balaústa.
Balaústa.


- Frutos simples esquizocárpicos
Por ejemplo:
"Esquizocarpo de mericarpios" y "carpóforo" en Terminología descriptiva de las plantas#Glosario de Apiaceae.
Un esquizocarpo de folículos es un fruto en el que (generalmente dos) carpelos de un pistilo se dividen a la madurez, cada carpelo desarrollándose en una unidad folículo, como en Asclepias.
Boraginaceae y Lamiaceae tienen un ovario que se vuelve 4-lobado durante el desarrollo, con el estilo de posición ginobásica, cada lóbulo se desarrolla en una núcula, que luego se separan a la madurez del fruto: sus frutos son por lo tanto un esquizocarpo de núculas. Las núculas esquizocárpicas típicas de estas dos familias llevaron a muchos autores a suponer que estaban cercanamente emparentadas, si bien pareciera que el desarrollo de su ovario y fruto fue independiente (Castellanos Cohen 2012[20]).
El esquizocarpo de aquenios de la familia Geraniaceae es llamado regma[13] procede de un gineceo en el cual todos los estilos se sueldan en un solo cuerpo. A la madurez y debido a tensiones higroscópicas, los carpelos se separan, cada uno con su estilo persistente, se abren, y catapultan las semillas, como en el geranio, Geranium.
Las aceráceas poseen un ovario 2-locular que desarrollará alas y dará como fruto un esquizocarpo de sámaras.
-
 Esquizocarpo de folículos en Asclepias, separados a la madurez.
Esquizocarpo de folículos en Asclepias, separados a la madurez.

-
 Flor de Boraginaceae, nótese el estilo ginobásico que divide al ovario en 4 lóbulos.
Flor de Boraginaceae, nótese el estilo ginobásico que divide al ovario en 4 lóbulos. -
 Los 4 lóbulos formarán 4 núculas en el esquizocarpo de núculas, fruto típico de Boraginaceae y también de Lamiaceae (las labiadas). Aquí se observan el estilo persistente y el cáliz persistente en el fruto.
Los 4 lóbulos formarán 4 núculas en el esquizocarpo de núculas, fruto típico de Boraginaceae y también de Lamiaceae (las labiadas). Aquí se observan el estilo persistente y el cáliz persistente en el fruto. -
 Flor de Geranium, ovario 4-lobado desarrollándose en fruto con estilo persistente, y a la derecha fruto maduro (regma) en el que dos semillas ya fueron catapultadas.
Flor de Geranium, ovario 4-lobado desarrollándose en fruto con estilo persistente, y a la derecha fruto maduro (regma) en el que dos semillas ya fueron catapultadas.


.jpg)
-
 Regma de Geranium sanguineum, donde ya se separaron las unidades de fruto y catapultaron las semillas.
Regma de Geranium sanguineum, donde ya se separaron las unidades de fruto y catapultaron las semillas. -
 El ovario de Acer (el maple) forma dos sámaras que se separan a la madurez, es un esquizocarpo de sámaras.
El ovario de Acer (el maple) forma dos sámaras que se separan a la madurez, es un esquizocarpo de sámaras. -
 Esquizocarpo de sámaras y unidad sámara en Acer pseudoplatanus.
Esquizocarpo de sámaras y unidad sámara en Acer pseudoplatanus.



Véase también[editar]
Referencias[editar]
- ↑ González, A.M. y Arbo, M.M. «Flor, simetría y sexualidad floral». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 17 de septiembre de 2009. Consultado el 10 de abril de 2009.
- ↑ González, A.M. y Arbo, M.M. «Simetría y sexualidad floral». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 17 de septiembre de 2009. Consultado el 12 de septiembre de 2012.
- ↑ González, A.M. y Arbo, M.M. «Androceo, número de estambres». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 14 de abril de 2009.
- ↑ Dimitri, M. 1987. Enciclopedia Argentina de Agricultura y Jardinería. Tomo I. Descripción de plantas cultivadas. Editorial ACME S.A.C.I., Buenos Aires.
- ↑ a b Error en la cita: Etiqueta
<ref>no válida; no se ha definido el contenido de las referencias llamadasUNNE gineceo ovario - ↑ Heslop Harrison y K. R. Shivanna. 1997. The receptive surface of the angiosperm stigma. Ann. Bot. 41: 1233-1258.
- ↑ a b Error en la cita: Etiqueta
<ref>no válida; no se ha definido el contenido de las referencias llamadasUNNE gineceo estilo - ↑ Error en la cita: Etiqueta
<ref>no válida; no se ha definido el contenido de las referencias llamadasSimpson 2005 - ↑ González, A.M. y Arbo, M.M. «Óvulo». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 10 de septiembre de 2012.
- ↑ Font Quer, P. (1982). Diccionario de Botánica. 8ª reimpresión. Barcelona: Editorial Labor, S. A. 84-335-5804-8.
- ↑ a b c d e f g h i Error en la cita: Etiqueta
<ref>no válida; no se ha definido el contenido de las referencias llamadasJudd et al. 2007 - ↑ González, A.M. y Arbo, M.M. «Clasificación de las Inflorescencias: Complejas simpodiales». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Consultado el 12 de septiembre de 2012.
- ↑ a b c d e f g h González, A.M. y Arbo, M.M. «Fruto». Morfología de Plantas Vasculares. Argentina: Universidad Nacional del Nordeste. Archivado desde el original el 21 de abril de 2015. Consultado el 12 de septiembre de 2012.
- ↑ van der Pijl, L. 1972. Principles of dispersal in higher plants. McGraw-Hill, Nueva York.
- ↑ Gray, A. 1877. Gray's lessons in botany and vegetable physiology. Ivison, Blackman, Taylor, Nueva York.
- ↑ Judd, W. S. 1985. A revised traditional/descriptive classification of fruits for use in floristics and teaching. Phytologia 58: 233-242.
- ↑ Error en la cita: Etiqueta
<ref>no válida; no se ha definido el contenido de las referencias llamadasWeberling 1989 - ↑ Spjut, R. W. 1994. A systematic treatment of fruit types. Mem. N.Y. Bot. Gard. 70: 1-182.
- ↑ 18-Poricidal capsule, "Poricidally dehiscent fruits use a "shaker" mechanism to disperse the seeds. The capsules are borne on long stalks, and wind causes the capsules to sway back and forth, shaking the seeds out through the pores." http://www.life.illinois.edu/help/digitalflowers/Fruits/18.htm
- ↑ Castellanos, Cohen. 2012. Comparative gynoecial development in Boraginaceae and Lamiaceae. Poster. http://2012.botanyconference.org/engine/search/index.php?func=detail&aid=789
| Control de autoridades |
|
|---|
 Datos: Q48789366
Datos: Q48789366