
Clorofila

Las clorofilas (del griego χλωρος, chloros, "verde", y φύλλον, fýlon, "hoja")[1] son una familia de pigmentos de color verde que se encuentran en las cianobacterias y en todos aquellos organismos que contienen cloroplastos o membranas tilacoidales en sus células, lo que incluye a las plantas y a las diversas algas. La clorofila es una biomolécula indispensable, crítica en la fotosíntesis, que es un proceso que permite a las plantas y algas almacenar energía a partir de la luz solar.
Historia
[editar]La clorofila fue descubierta en 1817 por los químicos franceses Pierre Pelletier (1788-1842) y Joseph Caventou (1795-1877), que consiguieron aislarla de las hojas de las plantas.[2] Pelletier introdujo los métodos, basados en la utilización de disolventes suaves, que permitieron por primera vez aislar no solo la clorofila, sino sustancias de gran importancia farmacológica como la cafeína, la colchicina o la quinina.
Descripción
[editar]Las clorofilas son un grupo de pigmentos que se encuentran en aquellos organismos eucariontes que poseen cloroplastos (plantas, algas) y en algunos procariotas: bacterias que no poseen cloroplastos (cianobacterias, bacterias verdes y púrpuras), y cuyos pigmentos se encuentran en sistemas de membrana internos: vesículas, lamelas y cromatóforos, pertenecientes a los dominios Eubacteria y Eucarya.
Estructura química de las moléculas de clorofila
[editar]
La estructura de las moléculas de clorofila tiene dos partes: un anillo de porfirina que contiene magnesio y cuya función es absorber luz, y una cadena hidrófoba de fitol cuya función es mantener la clorofila integrada en la membrana fotosintética.
Se conocen varias clorofilas. Todas se definen como derivados de la clorina original por la presencia de un quinto anillo que contiene cetona, además de los cuatro anillos pirrólicos. La mayoría de las clorofilas se clasifican como clorinas, que son parientes reducidas de las porfirinas (presentes en la hemoglobina). Comparten una vía biosintética común con las porfirinas, incluido el precursor uroporfirinógeno III. A diferencia de las heminas, que contienen hierro unido al centro N4, la mayoría de las clorofilas se unen al magnesio. Los ligandos axiales unidos al centro de Mg2+ se omiten a menudo para mayor claridad. Al anillo de clorina se le añaden varias cadenas laterales, que suelen incluir una larga cadena de fitilo (C20H39O). La forma más ampliamente distribuida en las plantas terrestres es la clorofila a. La única diferencia entre la clorofila a y la clorofila b es que la primera tiene un grupo alquilo mientras que la segunda tiene un grupo aldehído. Esta diferencia provoca una diferencia considerable en el espectro de absorción, lo que permite a las plantas absorber una mayor parte de la luz visible.[3][4]
- Estructuras de las clorofilas
-
 Clorofila a
Clorofila a -
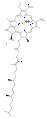 Clorofila b
Clorofila b -
 Clorofila c1
Clorofila c1 -
 Clorofila c2
Clorofila c2 -
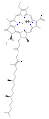 Clorofila d
Clorofila d -
 Clorofila f
Clorofila f


Localización en las células
[editar]Las clorofilas se encuentran en las membranas de los tilacoides, que en las cianobacterias son invaginaciones de la membrana plasmática, y en los plastos de las células eucarióticas son vesículas distribuidas por su interior. Las clorofilas aparecen insertas en la membrana, a las que se anclan por la cadena lateral constituida por un resto de fitol, asociadas a proteínas y otros pigmentos, con los que forman los fotosistemas.
Cada fotosistema contiene alrededor de 200 moléculas de clorofila, además de pigmentos auxiliares, con los que constituye la llamada antena. La antena está formada por conjuntos ordenados de moléculas de clorofila, otros pigmentos y proteínas, que se llaman complejos colectores de la luz. Solo una molécula de clorofila a en cada fotosistema convierte propiamente la energía radiante (luz) en energía química, cuando recibe un fotón con energía suficiente desde las moléculas de la antena, que se la van pasando.
Espectro de absorción y color
[editar]
Las clorofilas tienen típicamente dos tipos de absorción en el espectro visible, uno en el entorno de la luz azul (400-500 nm de longitud de onda), y otro en la zona roja del espectro (600-700 nm); sin embargo reflejan la parte media del espectro, la más nutrida y correspondiente al color verde (500-600 nm). Esta es la razón por la que las clorofilas tienen color verde y se lo confieren a los organismos, o a aquellos tejidos que tienen cloroplastos activos en sus células, así como a los paisajes que forman las plantas con clorofila.
Diversidad y distribución taxonómica
[editar]Las distintas formas de la clorofila se distribuyen desigualmente en la diversidad de los fotosintetizadores oxigénicos. La tabla siguiente presenta las diferentes formas de la clorofila y resumen su distribución sistemática.[5]
| Clorofila a | Clorofila b | Clorofila c1 | Clorofila c2 | Clorofila d | clorofila f | |
|---|---|---|---|---|---|---|
| Fórmula empírica | C55H72O5N4Mg | C55H70O6N4Mg | C35H30O5N4Mg | C35H28O5N4Mg | C54H70O6N4Mg | C55H70O6N4Mg |
| Grupo C2 | -CH3 | -CH3 | -CH3 | -CH3 | -CH3 | -CHO |
| Grupo C3 | -CH=CH2 | -CH=CH2 | -CH=CH2 | -CH=CH2 | -CHO | -CH=CH2 |
| Grupo C7 | -CH3 | -CHO | -CH3 | -CH3 | -CH3 | -CH3 |
| Grupo C8 | -CH2CH3 | -CH2CH3 | -CH2CH3 | -CH=CH2 | -CH2CH3 | -CH2CH3 |
| Grupo C17 | -CH2CH2COO-Phytyl | -CH2CH2COO-Phytyl | -CH=CHCOOH | -CH=CHCOOH | -CH2CH2COO-Phytyl | -CH2CH2COO-Phytyl |
| Enlace C17-C18 | Simple | Simple | Doble | Doble | Simple | Simple |
| Distribución | Universal (plantas y algas) | plantas y algas verdes | algas cromofitas | algas cromofitas | algunas cianobacterias | algunas cianobacterias |
- La clorofila a se encuentra en todos los casos, vinculada al centro activo de los complejos moleculares, llamados fotosistemas, que absorben la luz durante la fotosíntesis, y difiere de la clorofila b en que el radical de la posición 3 del grupo tetrapirrólico es -CH3 (metilo) en lugar de -CHO (grupo funcional de los aldehídos).
- La clorofila b caracteriza a los plastos de las algas verdes y de sus descendientes, las plantas terrestres (Viridiplantae). Esos plastos, y los organismos que los portan, son de color verde. También se encuentran plastos verdes en algunos grupos de protistas que han asimilado algas verdes unicelulares endosimbiontes adquiriendo así plastos secundarios. Se puede citar a las euglenas, a los cloraracniófitos y a algunos dinoflagelados, como Gymnodinium viride. También se encuentra en algunas cianobacterias (las cloroxibacterias), que por ello son de color verde planta en vez de azuladas; hace algún tiempo se les atribuyó por este rasgo el carácter de antepasados de los plastos verdes, pero luego se ha comprobado que es un carácter adquirido independientemente en varias líneas separadas.
- Las clorofilas c1 y c2 son características de un extenso y diverso clado de protistas que coincide con el superfilo Chromista y que incluye grupos tan importantes como las algas pardas, diatomeas, xantofíceas, haptófitas y criptófitas.[6]
- La clorofila d sólo se ha conocido durante decenios por una observación aislada y no repetida en un alga roja. Luego se ha encontrado en una cianobacteria (Acaryochloris marina), que parece especialmente apta para explotar luz roja cuando crece bajo ciertas ascidias. Recientemente se ha descubierto que esta clorofila no es propia del alga roja, sino que procede de la cianobacteria que vive epifita sobre estas algas.[7]
- La clorofila f ha sido encontrada en cianobacterias de estromatolitos de Australia.
También se encuentran clorofilas en animales que albergan dentro de sus células o entre ellas algas unicelulares (zooclorelas y zooxantelas). Gracias a esta simbiosis la fotosíntesis contribuye de manera significativa a la nutrición de corales, tridacnas, nudibranquios y otros animales marinos.
No todos los organismos fotosintetizadores tienen clorofilas. Las bacterias que no son cianobacterias tienen pigmentos muy distintos llamados bacterioclorofilas.
Biosíntesis
[editar]En algunas plantas, la clorofila deriva del glutamato y se sintetiza a lo largo de una vía biosintética ramificada que comparte con el heme y el siroheme.[8][9][10] La clorofila sintasa[11] es la enzima que completa la biosíntesis de la clorofila a:[12][13]
- clorofilida a + fitil difosfato clorofila a + difosfato

Esta conversión forma un éster del grupo ácido carboxílico del clorofílido a con el alcohol diterpénico de 20 carbonos fitol. La clorofila b es producida por la misma enzima que actúa sobre la clorofílida b. Lo mismo se sabe de la clorofila d y f, ambas producidas a partir de las correspondientes clorofílidas producidos en última instancia a partir de la clorofílida a.[14]
En las plantas angiospermas, los últimos pasos de la ruta biosintética dependen de la luz. Estas plantas son pálidas (etioladas) si crecen en la oscuridad. Las plantas no vasculares y las algas verdes tienen una enzima adicional independiente de la luz y crecen verdes incluso en la oscuridad[15]
La clorofila está unida a proteínas. La protoclorofilida, uno de los intermediarios biosintéticos, se presenta principalmente en forma libre y, en condiciones de luz, actúa como fotosensibilizador, formando radicales libres que pueden ser tóxicos para la planta. Por ello, las plantas regulan la cantidad de este precursor de la clorofila. En las angiospermas, esta regulación se consigue en el paso del ácido aminolevulínico (ALA), uno de los compuestos intermedios de la vía de biosíntesis. Las plantas que se alimentan de ALA acumulan niveles altos y tóxicos de protoclorofilida; lo mismo ocurre con los mutantes con un sistema regulador dañado.[16]
Medición del contenido de clorofila
[editar]
La medición de absorción de luz es compleja debido al solvente usado para extraer la clorofila de la planta, pues afecta los valores obtenidos.
- En éter etílico, la clorofila a tiene una absorbancia máxima aproximadamente entre los 430 nm y 662 nm, mientras la clorofila b tiene una absorbancia máxima entre los 453 nm y 642 nm.[17]
- El valor máximo de absorción de la clorofila a es entre los 465 nm y 665 nm. La clorofila a fluoresce a los 673 nm (máximo) y 726 nm. El valor máximo de absortividad de la clorofila a excede los 105 M−1 cm−1, que se encuentra entre los más altos para compuestos orgánicos de molécula pequeña.[18]
- En una concentración de 90 % de acetona-agua, la longitud de onda del máximo de absorción de la clorofila a son 430 nm y 664 nm; los máximos de la clorofila b son 460 nm y 647 nm; los máximos de la clorofila c1 son 442 nm y 630 nm; los máximos para la clorofila c2 son 444 nm y 630 nm; los máximos para la clorofila d son 401 nm, 455 nm y 696 nm.[19]
Midiendo la absorción de la luz en las regiones del rojo y rojo lejano, es posible estimar la concentración de clorofila que contiene una hoja.[20]
El coeficiente de fluorescencia se puede usar para medir el contenido de clorofila. Al excitar la clorofila, esta fluoresce a una longitud de onda más baja; la relación de emisión de fluorescencia a 705 ± 10 nm y 735 ± 10 nm puede proporcionar una relación lineal del contenido de clorofila comparable con las pruebas químicas. La relación entre F735/F700 provee un coeficiente de correlación r2 de 0,96 comparado con pruebas químicas, en el rango de 41 mg m−2 hasta 675 mg m−2. Gitelson desarrolló una fórmula para la lectura directa del contenido de clorofila en mg m−2. La fórmula proporcionó un método confiable para medir el contenido de clorofila desde 41 mg m−2 hasta 675 mg m−2 con un valor de correlación r2 de 0,95.[21]
Ecología
[editar]La clorofila puede detectarse fácilmente gracias a su comportamiento frente a la luz. Medir ópticamente la concentración de clorofila en una muestra de agua es sencillo y permite una estimación suficiente de la concentración de fitoplancton (algas microscópicas) e, indirectamente, de la actividad biológica; de esta manera la medición de clorofila es un instrumento importante de vigilancia de los procesos de eutrofización.
La presencia de clorofila es también medida por sistemas de teledetección, que informan sobre la distribución de la producción primaria, incluidas las oscilaciones estacionales y las fluctuaciones interanuales. De esta forma, la medición de la clorofila ayuda a la investigación del cambio climático y ecológico a escala global.
Véase también
[editar]Referencias
[editar]- ↑ «chlorophyll». Online Etymology Dictionary.
- ↑ Delépine, Marcel (septiembre de 1951). «Joseph Pelletier and Joseph Caventou». Journal of Chemical Education 28 (9): 454. Bibcode:1951JChEd..28..454D. doi:10.1021/ed028p454.
- ↑ Scheer H (2006). «An Overview of Chlorophylls and Bacteriochlorophylls: Biochemistry, Biophysics, Functions and Applications». Chlorophylls and Bacteriochlorophylls. Advances in Photosynthesis and Respiration (en inglés) 25. pp. 1-26. ISBN 978-1-4020-4515-8. doi:10.1007/1-4020-4516-6_1.
- ↑ Taniguchi M, Lindsey JS (January 2017). «Synthetic Chlorins, Possible Surrogates for Chlorophylls, Prepared by Derivatization of Porphyrins». Chemical Reviews (en inglés) 117 (2): 344-535. OSTI 1534468. PMID 27498781. doi:10.1021/acs.chemrev.5b00696.
- ↑ Chlorophyll molecules are specifically arranged in and around photosystems that are embedded in the thylakoid membranes of chloroplasts. Two types of chlorophyll exist in the photosystems: chlorophyll a and b.Speer, Brian R. (1997). «Photosynthetic Pigments». UCMP Glossary (online). University of California Museum of Paleontology. Consultado el 17 de julio de 2010.
- ↑ S. W. Jeffrey 1976. The occurrence of chlorophyll c1 and c2 in algae Journal of Phycology. Volume 12, Issue 3, pages 349-354
- ↑ Larkum, Anthony WD, and Michael Kühl. Chlorophyll d: the puzzle resolved. Trends in plant science 10.8 (2005): 355-357.
- ↑ Battersby AR (December 2000). «Tetrapyrroles: the pigments of life». Natural Product Reports 17 (6): 507-26. PMID 11152419. doi:10.1039/B002635M.
- ↑ Akhtar M (2007). «The Modification of Acetate and Propionate Side Chains During the Biosynthesis of Haem and Chlorophylls: Mechanistic and Stereochemical Studies». Ciba Foundation Symposium 180 - the Biosynthesis of the Tetrapyrrole Pigments. Novartis Foundation Symposia 180. pp. 131-155. ISBN 9780470514535. PMID 7842850. doi:10.1002/9780470514535.ch8.
- ↑ Willows RD (June 2003). «Biosynthesis of chlorophylls from protoporphyrin IX». Natural Product Reports 20 (3): 327-41. PMID 12828371. doi:10.1039/B110549N.
- ↑ Schmid HC, Rassadina V, Oster U, Schoch S, Rüdiger W (November 2002). «Pre-loading of chlorophyll synthase with tetraprenyl diphosphate is an obligatory step in chlorophyll biosynthesis». Biological Chemistry 383 (11): 1769-78. PMID 12530542. S2CID 3099209. doi:10.1515/BC.2002.198.
- ↑ Eckhardt U, Grimm B, Hörtensteiner S (September 2004). «Recent advances in chlorophyll biosynthesis and breakdown in higher plants». Plant Molecular Biology 56 (1): 1-14. PMID 15604725. S2CID 21174896. doi:10.1007/s11103-004-2331-3.
- ↑ Bollivar DW (November 2006). «Recent advances in chlorophyll biosynthesis». Photosynthesis Research 90 (2): 173-94. PMID 17370354. S2CID 23808539. doi:10.1007/s11120-006-9076-6.
- ↑ Tsuzuki Y, Tsukatani Y, Yamakawa H, Itoh S, Fujita Y, Yamamoto H (March 2022). «Effects of Light and Oxygen on Chlorophyll d Biosynthesis in a Marine Cyanobacterium Acaryochloris marina». Plants 11 (7): 915. PMC 9003380. PMID 35406896. doi:10.3390/plants11070915.
- ↑ Muraki N, Nomata J, Ebata K, Mizoguchi T, Shiba T, Tamiaki H, Kurisu G, Fujita Y (May 2010). «X-ray crystal structure of the light-independent protochlorophyllide reductase». Nature 465 (7294): 110-4. Bibcode:2010Natur.465..110M. PMID 20400946. S2CID 4427639. doi:10.1038/nature08950.
- ↑ Meskauskiene R, Nater M, Goslings D, Kessler F, op den Camp R, Apel K (October 2001). «FLU: a negative regulator of chlorophyll biosynthesis in Arabidopsis thaliana». Proceedings of the National Academy of Sciences of the United States of America 98 (22): 12826-31. Bibcode:2001PNAS...9812826M. JSTOR 3056990. PMC 60138. PMID 11606728. doi:10.1073/pnas.221252798.
- ↑ Gross, Jeana (1991). Pigments in vegetables: chlorophylls and carotenoids. Van Nostrand Reinhold. ISBN 978-0442006570.
- ↑ «Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy». Biochimica et Biophysica Acta (BBA) - Bioenergetics 975 (3): 384-394. 1989. doi:10.1016/S0005-2728(89)80347-0.
- ↑ Photosynthesis in algae. London: Kluwer. 2003. ISBN 978-0-7923-6333-0.
- ↑ «Joseph Pelletier and Joseph Caventou». Journal of Tree Physiology 23 (15): 1077-1079. September 2003. PMID 12975132. doi:10.1093/treephys/23.15.1077.
- ↑ «The Chlorophyll Fluorescence Ratio F735/F700 as an Accurate Measure of Chlorophyll Content in Plants». Remote Sens. Enviro. 69 (3): 296-302. 1999. Bibcode:1999RSEnv..69..296G. doi:10.1016/S0034-4257(99)00023-1.
Enlaces externos
[editar] Wikimedia Commons alberga una categoría multimedia sobre Clorofila.
Wikimedia Commons alberga una categoría multimedia sobre Clorofila.
| Control de autoridades |
|
|---|
 Datos: Q43177
Datos: Q43177- Multimedia: Chlorophyll / Q43177