
Foraminifera
| Foraminíferos | ||
|---|---|---|
| Rango temporal: 630–0Ma Ediacárico - Reciente | ||
 Ammonia tepida | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| (sin rango) | Supergrupo SAR | |
| Superfilo: | Rhizaria | |
| Filo: |
Foraminifera d'Orbigny, 1826 | |
| Clases y órdenes | ||
| ||
Los foraminíferos (Foraminifera, latín "portadores de orificios"[1]) son protistas ameboides, principalmente marinos, aunque también hay especies de agua dulce, caracterizados por un esqueleto o concha constituido por una o más cámaras interconectadas que fosiliza con relativa facilidad. Son llamados a veces coloquialmente "arena viviente", en alusión a su hábitat bentónico en el fondo marino mimetizándose con la arena. Los septos que separan las cámaras están perforados por uno o más orificios de interconexión denominados forámenes, los cuales dan nombre al grupo. Su protoplasma está diferenciado en un endoplasma y un ectoplasma del cual emergen seudópodos retráctiles (en este caso, reticulopodios granulados) que el organismo usa para la locomoción, captura de presas y creación de su esqueleto calcáreo.
Los foraminíferos suelen considerarse como el grupo más importante de microfósiles marinos debido a que son organismos muy abundantes en los sedimentos. Además, presentan una gran diversidad de especies, que son de gran utilidad en los estudios de tipo bioestratigráfico, paleoecológico, paleoceanográfico, etc.[2]
Se han descrito unas 10 000 especies actuales de foraminíferos,[3][4] además de unas 40 000 especies extintas, conocidas por sus fósiles.[5] Por lo general tienen un tamaño menor a 1 mm, pero algunas especies alcanzan hasta 20 cm.[6]
Características[editar]
Ecología[editar]
Los foraminíferos son fundamentalmente marinos y la mayoría viven sobre o dentro de los sedimentos del fondo marino (es decir, son bentónicos), mientras que algunos flotan en la columna de agua a distintas profundidades (es decir, son planctónicos). Unos pocos se han encontrado en agua dulce o en condiciones salobres y se han identificado algunas especies en el suelo a través del análisis molecular de ADN ribosómico.[7][8] Se los encuentra en todas las latitudes y profundidades, incluyendo las regiones polares y las fosas abisales.
El tamaño típico de los foraminíferos está comprendido entre 0,1 y 0,5 cm, aunque existen especies con tamaños que van desde 100 µm hasta casi 20 cm. Los foraminíferos son organismos heterótrofos que se alimentan de pequeños organismos tales como diatomeas o bacterias que capturan con una red de seudópodos. Las especies de foraminíferos más grandes son capaces de capturar nematodos y crustáceos.
Algunos foraminíferos tienen como endosimbiontes a algas unicelulares de diversos linajes, tales como algas verdes, algas rojas, algas doradas, diatomeas y dinoflagelados. Las especies más grandes pueden contener una gran cantidad de algas, densamente empaquetadas, constituyendo verdaderas granjas de algas. Ciertos foraminíferos son cleptoplásticos, es decir, conservan los cloroplastos de las algas que ingieren, de forma que estos orgánulos siguen realizando la fotosíntesis dentro del foraminífero.[9]




Granurreticulopodios[editar]
Las dos características distintivas de los foraminíferos son la concha y la red de seudópodos. La célula de los foraminíferos se divide en un endoplasma granular situado dentro de la concha y en un ectoplasma transparente que emerge de la concha a través de una abertura principal o de muchas perforaciones. El ectoplasma usualmente rodea completamente la concha y es el responsable de su construcción, apoyado por los seudópodos. Existe un flujo continuo de citoplasma entre las cámaras de la concha, a través de los orificios internos, incluso en las especies con complejos caparazones.
Los seudópodos se forman a partir del ectoplasma y están limitados por la membrana celular y sostenidos por microtúbulos. Los foraminiferos presentan mecanismos para el rápido ensamblado y desensamblado de los microtúbulos, que permiten la rápida extensión y retracción de los seudópodos. Estos son utilizados para la locomoción, fijación al sustrato, construcción de la concha y para la captura de alimento. La longitud de los seudópodos depende de la especie: en algunas los seudópodos son dos o tres veces la longitud del cuerpo, mientras que en otras alcanzan hasta 20 veces esta longitud.[10] El movimiento de los foraminíferos se produce mediante la fijación de algunos seudópodos al sustrato, empujando al resto de la célula. La velocidad típica de desplazamiento es de 1 a 2,5 cm/hora.
Los seudópodos se denominan granurreticulopodios (reticulopodios granulados), pues están constituidos por citoplasma granular, y son largos, finos y ramificados, formando por amontonamiento (anastomosis) la red de reticulopodios. A lo largo de los seudópodos se produce un flujo citoplasmático bidireccional que lleva gránulos, los cuales pueden consistir en partículas de diversos materiales, mitocondrias, vacuolas digestivas o que contienen productos de desecho, dinoflagelados simbióticos, etc. Por esta razón uno de los sinónimos del grupo es Granuloreticulosa, aunque el proceso de transporte de los gránulos todavía no se comprende completamente.
Esqueleto[editar]

La mayoría de las especies forman un esqueleto intraectoplásmico denominado concha, testa o caparazón. Es la característica más sobresaliente de los foraminíferos y el motivo de que sean susceptibles de fosilizar con relativa facilidad. Solo son monotálamos (esqueleto de una sola cámara) los foraminíferos primitivos. La mayoría de los foraminíferos son politálamos, es decir, su concha consta de varias cámaras. Estas están separadas por septos que presentan uno o más orificios de interconexión denominados forámenes que, además, dan el nombre al grupo. El interior de las cámaras se encuentra forrado por una película orgánica de naturaleza próxima a la quitina.[11][12][13] La concha presenta una o más aberturas principales y además puede presentar pequeños poros al exterior (conchas perforadas) o carecer de ellos (conchas imperforadas). Los seudópodos salen al exterior a través de las aberturas principales y en el caso de las conchas perforadas también a través de los poros cuando estos son lo suficientemente anchos.
Cuando existe más de una cámara, la primera que se forma se denomina proloculus y es generalmente la más pequeña. Las cámaras están separados por septos cuya intersección con la concha produce una línea de contacto o sutura. Los orificios o forámenes presentes en los septos permiten el paso de alimentos, vacuolas contráctiles y núcleos. Las aberturas presentes en la pared de la última cámara conectan el endoplasma interno con los seudópodos y permiten la liberación de las células hijas.[14]
La concha puede tener paredes de material orgánico, aglutinado (formadas por partículas recogidas por el organismo y cementadas) o mineralizado (formadas por sustancias minerales secretadas por el organismo). En las especies desnudas (formas ameboides sin concha, Athalamea) el cuerpo celular está constituido por una sola cámara delimitada por venas gruesas embebidas en un material mucoide y amorfo. Las formas con concha más primitivas (por ejemplo, Allogromia) presentan una pared delgada compuesta por material proteínico con poco o ningún material aglutinado. La transición entre las conchas orgánicas y las aglutinadas tiene lugar en varios linajes de foraminíferos monotálamos (Monothalamea). En las especies con paredes aglutinadas, las partículas sedimentarias recogidas y cementadas por el organismo limitan una matriz orgánica amorfa o fibrosa. Por último, en las formas con múltiples cámaras o politálamos (Tubothalamea y Globothalamea), las paredes contienen material aglutinado o mineralizado, incluyendo calcita, aragonita y sílice.
Núcleo y orgánulos celulares[editar]
El endoplasma de los foraminíferos monotálamos es homogéneo, con los orgánulos celulares distribuidos uniformemente. En los foraminíferos politálamos las primeras cámaras constituyen el segmento cromático, conteniendo el núcleo y otros orgánulos, mientras que últimas cámaras constituyen los segmentos vegetativos y son responsables de la digestión del alimento. La célula de algunas especies presenta un único núcleo, que puede alcanzar un tamaño bastante grande, de hasta 100 µm de diámetro, mientras que otras especies contienen cientos de pequeños núcleos dispersos por el citoplasma. En los foraminiferos con alternancia de generaciones, el número de núcleos depende de la generación: los gamontes son usualmente uninucleares, mientras que los esquizontes son plurinucleados. En algunos foraminiferos existen dos tipos de núcleos: el único núcleo somático (que participa solo en las funciones metabólicas) es de tamaño grande y ocupa las cámaras periféricas, mientras que los núcleos generativos (que participan en la formación de gametos) son numerosos, pequeños y ocupan la parte central de la concha.
Además del núcleo, los elementos dominantes presentes en el citoplasma son las grandes vacuolas digestivas. Estas se forman en el exterior de la concha, englobando el alimento, aunque la digestión ocurre principalmente en el interior. El aparato de Golgi es similar al de los otros protistas. Tanto el ectoplasma como el endoplasma contienen numerosas pequeñas mitocondrias que presentan las crestas mitocondriales tubulares típicas de muchos protistas.[15] Otros orgánulos celulares presentes en las células de los foraminíferos son los microsomas y los peroxisomas, utilizados en el metabolismo y en la degradación de las sustancias tóxicas.
Ciclo de vida[editar]
El ciclo de la vida de los foraminíferos comprende a menudo una alternancia de generaciones con reproducción asexual y sexual. La alternancia generacional se traduce en dos morfologías diferentes (dimorfismo), que a simple vista son muy similares.[16][17] Así, en el ciclo reproductivo, existen dos generaciones diferentes:
- Gamonte. De tipo haploide, uninucleado, microsférico (tamaño pequeño) y con reproducción sexual mediante gamogonia.
- Esquizonte. De tipo diploide, plurinucleado, megalosférico (o macrosférico, tamaño grande) y con reproducción asexual mediante esquizogonia.
El gamonte inicialmente presenta un solo núcleo y se divide para producir numerosos gametos, que normalmente tienen dos flagelos. El esquizonte es multinucleado, y después de la meiosis se fragmenta para producir nuevos gamontes. No es infrecuente que haya varias rondas de reproducción asexual por cada una de generación sexual entre las formas bentónicas. De hecho, entre las poblaciones actuales la proporción de esquizontes es mucho mayor que la de gamontes, lo que indica que la reproducción sexual es más excepcional. La forma microsférica es conservadora en el sentido ontogenético, pues recapitula el desarrollo filogenético del grupo. Por el contrario, la forma megalosférica es progresiva, al comenzar directamente su desarrollo en formas evolutivamente avanzadas.
Tipos de foraminíferos[editar]


Los foraminíferos presentan dos modos de vida claramente diferenciados: bentónico y planctónico, viviendo en el sedimento o en la columna de agua.[18] Los foraminíferos son utilizados como indicadores de la profundidad de la columna de agua, la temperatura, productividad,[19] volumen total de hielo así como características geoquímica del agua (pH, concentración de metales trazas), y contaminación del medio.
Foraminíferos bentónicos[editar]
Los foraminíferos bentónicos, cuyos primeros fósiles datan de finales Precámbrico, pueden ser tanto sésiles como vágiles. Según la posición que ocupan en el sedimento se pueden clasificar en epibentónicos o endobentónicos (epifaunales o infaunales, respectivamente). Dos parámetros ambientales son los principales responsables en determinar la profundidad en la que un foraminífero vive y su abundancia, la disponibilidad de oxígeno y la cantidad de alimento disponible. Se valen de sus seudópodos o de secreciones calcáreas para fijarse al sustrato. La mayoría son marinos y estenohalinos (esto es, toleran pequeñas variaciones en los niveles de salinidad de las aguas). Algunos foraminíferos con conchas de tipo aporcelanado viven en ambientes hipersalinos (salinidad > 35 ‰), mientras que algunos foraminíferos aglutinados o hialinos prefieren aguas salobres, incluso algunos son capaces de adaptarse a las más variadas condiciones de salinidad (salvo lagos).
Foraminíferos planctónicos[editar]
Los foraminíferos planctónicos son mucho menos diversos que los bentónicos (con alrededor de 50 especies modernas) además de presentar un menor registro geológico, ya que los primeros fósiles son del período Jurásico. Son típicamente estenohalinos (salinidad entre 34 y 36 ‰). Suelen ocupar la zona fótica, y durante su ciclo vital algunas especies varían de profundidad (migración ontogénica). La profundidad límite suele ser alrededor de 300 m, y frecuentemente la mayor parte de las especies suele habitar a profundidades inferiores a los 50 m.
Las biocenosis de foraminíferos presentan una mayor diversidad de especies y con morfologías más complejas e individuos de mayor tamaño en los ambientes tropicales y subtropicales (asociaciones politáxicas).
Por el contrario, en ambientes de alta latitud las asociaciones de foraminíferos planctónicos suelen ser poco diversas, con organismos de pequeño tamaño y formas sencillas, y suelen estar dominadas por una o a lo sumo dos especies (asociaciones oligotáxicas).
La concha[editar]

Tipos de conchas[editar]
Las conchas de los foraminíferos pueden estar constituidas por:
- Material orgánico (quitina).
- Partículas del medio ambiente unidas entre sí mediante un cemento (calcítico o quitinoideo) segregado por el organismo, llamadas conchas aglutinadas.
- Material calcáreo (calcita o menos frecuentemente aragonito) segregado enteramente por el organismo. Dependiendo de la disposición de los cristales, las conchas pueden ser aporcelanadas o hialinas.
- Muy raramente (un solo género) presentan conchas silíceas.
Las conchas aporcelanadas o "imperforadas" presentan cristales de muy pequeño tamaño (0,1-2 µm), con formas aciculares o globulares y dispuestos aleatoriamente. Al no existir paralelismo entre sus ejes ópticos, con el microscopio con luz transmitida la concha se ve negra y opaca, mientras que con luz reflejada se ve blanca y brillante. Las conchas aporcelanadas suelen tener seudoporos que no suelen atravesar la concha por completo.
Las conchas hialinas o "perforadas" se caracterizan por su transparencia, que es el resultado de la delgadez de las paredes en las conchas granulares, o de la disposición regular de los cristales con un eje óptico perpendicular a la superficie del concha en las fibroradiales. Existen también conchas hialinas de estructura lamelar, formadas por la superposición de laminas calcáreas separadas por una película de proteína. La estructura lamelar puede ser monolamelar o plurilamelar en función del número de láminas calcáreas por cada cámara. Las conchas hialinas presentan perforaciones con diámetro, densidad y localización variables.
Tradicionalmente la taxonomía en foraminíferos ha estado basada en la morfología y geometría de sus conchas o testas:
- Rasgos a nivel de familia: plano general de la disposición de las cámaras.
- Rasgos a nivel de género: forma general del caparazón, situación de la abertura, forma de las cámaras, construcción de la pared, carácter de los poros y de las espinas, concentración de poros.
- Rasgos a nivel de especie: número de cámaras, carácter de las suturas, detalles de las aberturas.
- Rasgos a nivel de variedad: detalles de la ornamentación que dependen de la variabilidad del medio.
Disposición de las cámaras[editar]

Las conchillas de los foraminíferos dan lugar a unos espacios llamados cámaras. El número, forma y la disposición de estas cámaras es uno de los criterios de clasificación básico en la taxonomía de estos organismos.
- Número de cámaras:
Las conchas pueden dar lugar a una única cámara, son las conchas uniloculares (foraminíferos monotálamos), o pueden estar formadas por varias cámaras, en este caso se trata de conchas pluriloculares (foraminíferos politálamos).
- Disposición de las cámaras:
Esta clasificación evidentemente hace referencia únicamente a las conchas pluriloculares. En el caso de las conchas uniloculares, el criterio de diferenciación es la forma de su única cámara (e.g., esférica, tubular, espiralada,…).
La disposición de las cámaras puede ser:
- Seriada, las cámaras se sitúan en relación longitudinal a un eje. Pueden ser uniloculares, biloculares, triloculares, o mixtas.
- Espiralada, esta es la denominación cuando las cámaras sufren un proceso de enrollamiento según una espiral. Este enrollamiento puede ser en un único plano, son las conchas planispiraladas, que a su vez pueden ser involutas o evolutas dependiendo de si las cámaras son abrazadoras o no. El enrollamiento también se puede producir en un eje, con lo que se obtiene un diseño helicoidal, son las conchas trocoespiraladas.
- Agatistega, en este caso la disposición de las conchas se hace alternativamente según varios ejes.
- Ciclostega, se trata de cámaras circulares que se sitúan en un solo plano. El resultado son conchas planas o biconvexas de periferia circular.
- Orbitostega, son conchas con un crecimiento ciclostego al que se superpone otro de cámaras en un eje perpendicular al plano principal de crecimiento. El resultado son conchas de contorno leticular.
- Otras características de las conchas:
Además de la morfología de la concha es interesante observa la forma, posición y número de otros elementos morfológicos. Estos son la abertura principal de la concha, aberturas secundarias o accesorias, presencia de espinas, dientes, tegillas, bullas, costillas, estrías, carenas, murica, etc. Otras características interesantes son la estructura y textura superficial de la concha, la presencia o no de cuello, las suturas intercamerales, etc.
Clasificación[editar]
-1,_Rotalia_veneta_(M_Sch)-2,_Cornuspira_planorbis_(MSch)-3.jpeg)
La clasificación de los foraminíferos todavía no está resuelta, por lo que siendo conservadores podemos distinguir cinco clases:
- Athalamea. Incluye a las formas ameboides sin concha, probablemente el grupo ancestral de los foraminíferos.
- Monothalamea. Son un grupo de foraminíferos bentónicos que presentan conchas con una sola cámara (monocamerados o monotálamos). La pared de la concha es orgánica o aglutinada.
- Xenophyophorea. Es un grupo de foraminíferos especializados, bentónicos y detritívoros. Son multinucleados y gigantes para ser protistas, pues miden desde unos mm hasta 20 cm. El delicado esqueleto está constituido un sistema de tubos ramificados compuesto por sedimentos aglutinados.
- Tubothalamea. Son un grupo de foraminíferos bentónicos que presentan conchas bicameradas o multicameradas con cámaras tubulares al menos en el estadio juvenil, muchas veces enrolladas espiralmente. La pared de la concha es aglutinada o calcárea.
- Globothalamea. Son un grupo de foraminíferos bentónicos y planctónicos que presentan conchas multicameradas típicamente globulares (trocospiraladas, pero también triseriadas, biseriadas y uniseriadas). La pared de la concha es aglutinada o calcárea.
Los foraminíferos fueron subdivididos inicialmente en tres clases: Athalamea, Monothalamea y Polythalamea,[20][21] pero la última clase ha sido reciente dividida en dos: Tubothalamea y Globothalamea, mediante un estudio de filogenia molecular.[22] En las clasificaciones más recientes, Athalamea y Xenophyophorea se incluyen en Monothalamea, pero con la indicación de que este grupo es parafilético y que son necesarios estudios adicionales.[22][23]
La posición taxonómica y las relaciones filogenéticas de Athalamea todavía no están esclarecidas, de manera que algunas veces han sido consideradas como un grupo diferente del de los foraminíferos y otras veces como un grupo integrante, aunque ancestral, de los mismos. Este grupo, que incluye al orden Reticulomyxida, es por el momento considerado como incertae sedis al igual que los xenofióforos (clase Xenophyophorea) y los komokioideos (superfamilia Komokioidea y/o orden Komokiida).[24][25]
Importancia[editar]
Los foraminíferos son utilizados como bioindicadores de temperatura, salinidad, oxígeno, turbidez, e igualmente sus fósiles son importantes en interpretaciones paleoceanográficas y paleoclimáticas, ya que los cambios en las poblaciones de estos organismos indican modificaciones en el ambiente.
Por su pequeño tamaño, amplia distribución en mares y océanos, elevada biodiversidad y alta tasa de cambio evolutivo, sus fósiles son muy utilizados en bioestratigrafía para establecer biozonas muy detalladas, y así datar y correlacionar rocas procedentes de sondeos, lo cual les añade una importancia económica relacionada con la búsqueda de hidrocarburos.
Árbol filogenético[editar]
Las relaciones son aproximadamente las siguientes:[26]
| Foraminifera |
| |||||||||||||||||||||
Galería[editar]
-
Reticulomyxa filosa, sin concha (Athalamea)
-
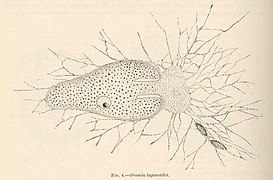 Allogromia lagenoides, concha monocámara (Monothalamea)
Allogromia lagenoides, concha monocámara (Monothalamea) -
 Xenofióforo (Xenophyophorea)
Xenofióforo (Xenophyophorea) -
 Amphisorus hemprichii, concha tubular espiralada (Tubothalamea)
Amphisorus hemprichii, concha tubular espiralada (Tubothalamea) -
 Globigerina, célula y concha globular (Globothalamea)
Globigerina, célula y concha globular (Globothalamea)




Véase también[editar]
- Foraminíferos bentónicos
- Foraminíferos planctónicos
- Clasificación de foraminíferos
- Barranco Hondo (Lorca)
Referencias[editar]
- ↑ Diccionario etimológico - Foraminífero
- ↑ Molina, E. (2002, 2004). Micropaleontología. Prensas Universitarias de Zaragoza, Colección Textos Docentes, 93. 634 pp. ISBN 84-7733-619-9
- ↑ Ald, S.M. et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ↑ «World Foraminifera Database».
- ↑ Pawlowski, J., Lejzerowicz, F., & Esling, P. (2014). Next-generation environmental diversity surveys of foraminifera: preparing the future. The Biological Bulletin, 227(2), 93-106.
- ↑ Marshall M (3 de febrero de 2010). «Zoologger: 'Living beach ball' is giant single cell». New Scientist.
- ↑ Giere, Olav (2009). Meiobenthology: the microscopic motile fauna of aquatic sediments (2nd edición). Berlin: Springer. ISBN 978-3540686576.
- ↑ Lejzerowicz, Franck; Pawlowski, Jan; Fraissinet-Tachet, Laurence; Marmeisse, Roland (1 de septiembre de 2010). «Molecular evidence for widespread occurrence of Foraminifera in soils». Environmental Microbiology 12 (9): 2518-26. PMID 20406290. doi:10.1111/j.1462-2920.2010.02225.x.
- ↑ Bernhard, J. M.; Bowser, S.M. (1999). «Benthic Foraminifera of dysoxic sediments: chloroplast sequestration and functional morphology». Earth Science Reviews 46: 149-165. Bibcode:1999ESRv...46..149B. doi:10.1016/S0012-8252(99)00017-3.
- ↑ Gupta, B.K.S. (Ed.) (1999). Modern foraminifera. Kluwer Academic Publishers.
- ↑ Hottinger, L. C. (1986). «Capítulo 14. Construction, structure, and function of foraminiferal shells». En: Leadbeater, S. C. y Riding, R. (eds.) Biomineralization in Lower Plants and Animals. The Systematics Association, Special Volume 30: 220-235
- ↑ Hemleben, C.; Roger-Anderson, O.; Berthold, W. y Spincler, M. (1986). «Capítulo 15. Calcification and chamber formation in Foraminifera - a brief overview». En: Leadbeater, S. C. y Riding, R. (eds.) Biomineralization in Lower Plants and Animals. The Systematics Association, Special Volume 30: 238-249.
- ↑ Anderson, O.R. y Lee, J. J. (1991). «Capítulo 2. Citology and fine structure». En: Lee, J. J. y Anderson, O. R. (eds.) Biology of Foraminifera. Academic Press, Harcourt Brace Jovanovich Publishers, Londres, 7-40
- ↑ Becker, S. Modern foraminifera: biological and ecological basics Archivado el 26 de septiembre de 2011 en Wayback Machine..
- ↑ Schaechter, M. (2009). Encyclopedia of microbiology. Academic Press.
- ↑ Loeblich Jr, A.R.; Tappan, H. (1964). «Foraminiferida». Part C, Protista 2. Treatise on Invertebrate Paleontology. Geological Society of America. pp. C55-C786. ISBN 0-8137-3003-1.
- ↑ Moore, R.C.; Lalicker, A.G.; Fischer, C.G. (1952). «Ch 2 Foraminifera and Radiolaria». Invertebrate Fossils. McGraw-Hill. OCLC 547380.
- ↑ Murray, J. W. (1991). «Ecology and distribution of planktonic foraminifera». En: Lee, J. J. y Anderson, O. R. (eds.) Biology of Foraminifera. Academic Press, Harcourt Brace Jovanovich Publishers, Londres, 221-284.
- ↑ Pardo, A. y Camara, N. (2007). «Foraminíferos, los legionarios del océano invisible». Inmersión, 88: 22-29
- ↑ Levine, N.D., Corliss J.O., Cox F.E.G., Deroux, G., Grain, J., Honigberg, B.M., Leedale, G.F., Loeblich III, A.R., Lom, J., Lynn, D., Merinfeld, E.G., Page, F.C., Poljansky, G., Sprague, V., Vavra, J. y Wallace, F.G. (1980). A newly revised classification of the Protozoa. Journal of Protozoology, 27: 37-58.
- ↑ Lee J.J., Hutner, S.H. y Bovee, E.C. (Eds.) (1985). An Illustrated Guide to the Protozoa. Society of Protozoologists, Allen, Lawrence, Kansas, 629 p.
- ↑ a b Pawłowski, J., Holzmann, M., y Tyszka, J. (2013). New supraordinal classification of Foraminifera: Molecules meet morphology Archivado el 8 de diciembre de 2015 en Wayback Machine.. Marine Micropaleontology, 100: 1–10.
- ↑ Adl, S.M. et al. (2012). The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ↑ Tendal, O.S. y Hessler, R.R. (1977). An introduction to the biology and systematics of the Komokiacea (Textulariina, Foraminiferida). Galathea Report, 14: 165-194.
- ↑ Tendal, O.S. (1996). Synoptic checklist and bibliography of the Xenophyophorea (Protista), with a zoogeographical survey of the group. Galathea 17: 79-102.
- ↑ Flakowski, 2005, Habura et al 2006, Longet & Pawlowski 2007, y Pawlowski 2003, graficado en http://palaeos.com/eukarya/rhizaria/foraminifera.html
Enlaces externos[editar]
 Wikimedia Commons alberga una categoría multimedia sobre Foraminifera.
Wikimedia Commons alberga una categoría multimedia sobre Foraminifera. Wikispecies tiene un artículo sobre Foraminifera.
Wikispecies tiene un artículo sobre Foraminifera.
- ZCO-1999. «Foraminíferos: los legionarios del océano invisible». Consultado el 19 de marzo de 2010.
- Foraminífero Galería - Catálogo ilustrado según género, localidad y era geológica
- Hottinger, Lukas (2006). «Illustrated glossary of terms used in foraminiferal research». Carnets de Géologie - Notebooks on Geology. Memoir 2006/02 (CG2006_M02). Brest. p. 126. Archivado desde el original el 16 de octubre de 2014. Consultado el 17 de noviembre de 2016.
| Control de autoridades |
|
|---|
 Datos: Q107027
Datos: Q107027- Multimedia: Foraminifera / Q107027
- Especies: Foraminifera