
TFIIH
| factor de transcripción general IIH, polipéptido 1, 62kDa | ||||
|---|---|---|---|---|
| Estructuras disponibles | ||||
| PDB | Buscar ortólogos: PDBe, RCSB | |||
| Identificadores | ||||
| Símbolos | GTF2H1 (HGNC: 4655) BTF2 | |||
| Identificadores externos | ||||
| Locus | Cr. 11 p15.1-p14 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
| factor de transcripción general IIH, polipéptido 2, 44kDa | ||||
|---|---|---|---|---|
| Estructuras disponibles | ||||
| PDB | Buscar ortólogos: PDBe, RCSB | |||
| Identificadores | ||||
| Símbolos | GTF2H2 (HGNC: 4656) BTF2, TFIIH, BTF2P44, T-BTF2P44 | |||
| Identificadores externos | ||||
| Locus | Cr. 5 q12.2-13.3 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
| factor de transcripción general IIH, polipéptido 3, 34kDa | ||||
|---|---|---|---|---|
| Estructuras disponibles | ||||
| PDB | Buscar ortólogos: PDBe, RCSB | |||
| Identificadores | ||||
| Símbolos | GTF2H3 (HGNC: 4657) BTF2, TFIIH | |||
| Identificadores externos | ||||
| Locus | Cr. 12 q24.31 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
El factor de transcripción general IIH, también conocido como factor de transcripción II humano o TFIIH, es un importante complejo proteico que desempeña funciones en la transcripción de varios genes que codifican proteínas y en los procesos de reparación por escisión de nucleótidos del ADN (NER). TFIIH fue descubierto en 1989 cuando el factor de transcripción general δ o factor de transcripción básico 2 se caracterizó como un factor de transcripción indispensable en ensayos in vitro. Este factor también se aisló de la levadura y finalmente se denominó TFIIH en 1992.[1][2]
TFIIH consta de diez subunidades, 7 de las cuales (ERCC2/XPD, ERCC3/XPB, GTF2H1/p62, GTF2H4/p52, GTF2H2/p44, GTF2H3/p34 y GTF2H5/TTDA) forman el complejo central. El subcomplejo quinasa activador de ciclina (CDK7, MAT1 y ciclina H) está vinculado al núcleo celular a través de la proteína XPD.[3] Dos de las subunidades, ERCC2/XPD y ERCC3/XPB, tienen actividades helicasa y ATPasa, respectivamente, y ayudan a crear el bucle de transcripción. En ensayos in vitro, estas subunidades solo se requieren para la transcripción si el ADN molde no está ya desnaturalizado o si está superenrollado.
Otras dos subunidades de TFIIH, CDK7 y ciclina H, fosforilan los aminoácidos de serina en el dominio C-terminal de la ARN polimerasa II y posiblemente otras proteínas involucradas en el ciclo celular. Además de una función vital en el inicio de la transcripción, TFIIH también participa en la reparación por escisión de nucleótidos.
Historia
[editar]Antes de que TFIIH fuera identificado, ya era conocido con otros nombres. Este factor se aisló por primera vez en 1989 de hígado de rata, bautizado en ese momento como factor de transcripción delta. También se aisló de una célula cancerosa conocida en ese momento como factor de transcripción básico 2, y se aisló de levadura con el nombre de factor de transcripción B. Finalmente, en 1992 fue rebautizado como TFIIH.[4]
Estructura
[editar]El TFIIH consta de dos subunidades principales: la que conforma el núcleo o XPB, que presenta las subunidades p62, p52, p44, p34 y p8; y CAK, compuesto por CDK7, ciclina H y MAT1. La unidad que une el núcleo XPB al CAK se llama XPD.[5]
Funciones
[editar]- Iniciación de la transcripción de genes codificantes.[6]
- Reparación de nucleótidos del ADN.[6]
TFIIH actúa para reclutador de la ARN polimerasa II hacia los promotores de los genes. Funciona como una helicasa que desenrolla el ADN. También desenrolla el ADN después de que una lesión de ADN ha sido reconocida por la vía de reparación del genoma global (GGR) o la vía de reparación por transcripción acoplada (TCR).[7][8] Además, el TFIIH purificado tiene un papel en la producción de ARN mediante la activación de la enzima alfa-amanitina.
Reparación del ADN
[editar]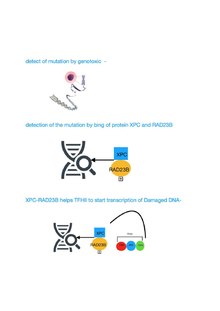
TFIIH participa en la reparación por escisión de nucleótidos (NER) abriendo la doble hélice de ADN después de que se reconoce inicialmente el daño. NER es una vía de varios pasos que elimina una amplia gama de diferentes daños que distorsionan el emparejamiento normal de bases, incluidos los daños químicos de gran calibre y los daños inducidos por rayos UV. Los individuos con defectos mutacionales en genes que especifican componentes proteicos que catalizan la vía NER, incluidos los componentes TFIIH, a menudo muestran patologías relacionadas con el envejecimiento prematuro.[9][10]
Inhibidores
[editar]Recientemente se han encontrado potentes productos naturales bioactivos como la triptolida que inhiben la transcripción en mamíferos mediante la inhibición de la subunidad XPB de TFIIH como un conjugado de glucosa para atacar las células cancerosas hipóxicas con una mayor expresión del transportador de glucosa.[11]
Patologías relacionadas
[editar]Se sabe que los polimorfismos genéticos de genes que codifican subunidades de TFIIH están asociados con una mayor susceptibilidad al cáncer en muchos tejidos, por ejemplo, el tejido cutáneo, el tejido mamario y el tejido pulmonar. Las mutaciones en las subunidades XPD o XPB pueden conducir a una variedad de enfermedades, como xeroderma pigmentosum (XP) o XP combinado con el síndrome de Cockayne.[12] Además de las variaciones genéticas, las proteínas codificadas por virus también se dirigen al TFIIH.[13] También cabe destacar que la mutación de los genes ERCC3/XPB, ERCC2/XPD o TTDA causa una patología denominada tricotiodistrofia, una afección caracterizada por fotosensibilidad, ictiosis, cabello y uñas quebradizos, deterioro intelectual, disminución de la fertilidad y/o baja estatura.[9]
Véase también
[editar]Referencias
[editar]- ↑ Flores O, Lu H, Reinberg D (February 1992). «Factors involved in specific transcription by mammalian RNA polymerase II. Identification and characterization of factor IIH». The Journal of Biological Chemistry 267 (4): 2786-93. PMID 1733973.
- ↑ Kim TK, Ebright RH, Reinberg D (May 2000). «Mechanism of ATP-dependent promoter melting by transcription factor IIH». Science 288 (5470): 1418-22. Bibcode:2000Sci...288.1418K. PMID 10827951. doi:10.1126/science.288.5470.1418.
- ↑ Lee TI, Young RA (2000). «Transcription of eukaryotic protein-coding genes». Annual Review of Genetics 34: 77-137. PMID 11092823. doi:10.1146/annurev.genet.34.1.77.
- ↑ Rimel JK, Taatjes DJ (June 2018). «The essential and multifunctional TFIIH complex». Protein Science 27 (6): 1018-1037. PMC 5980561. PMID 29664212. doi:10.1002/pro.3424.
- ↑ Drapkin R, Reardon JT, Ansari A, Huang JC, Zawel L, Ahn K, Sancar A, Reinberg D (April 1994). «Dual role of TFIIH in DNA excision repair and in transcription by RNA polymerase II». Nature 368 (6473): 769-72. Bibcode:1994Natur.368..769D. PMID 8152490. doi:10.1038/368769a0.
- ↑ a b Compe E, Egly JM (May 2012). «TFIIH: when transcription met DNA repair». Nature Reviews. Molecular Cell Biology 13 (6): 343-54. PMID 22572993. doi:10.1038/nrm3350.
- ↑ Hoogstraten D, Nigg AL, Heath H, Mullenders LH, van Driel R, Hoeijmakers JH, Vermeulen W, Houtsmuller AB (November 2002). «Rapid switching of TFIIH between RNA polymerase I and II transcription and DNA repair in vivo». Molecular Cell 10 (5): 1163-74. PMID 12453423. doi:10.1016/s1097-2765(02)00709-8.
- ↑ Assfalg R, Lebedev A, Gonzalez OG, Schelling A, Koch S, Iben S (January 2012). «TFIIH is an elongation factor of RNA polymerase I». Nucleic Acids Research 40 (2): 650-9. PMC 3258137. PMID 21965540. doi:10.1093/nar/gkr746.
- ↑ a b Theil AF, Hoeijmakers JH, Vermeulen W (November 2014). «TTDA: big impact of a small protein». Experimental Cell Research 329 (1): 61-8. PMID 25016283. doi:10.1016/j.yexcr.2014.07.008.
- ↑ Edifizi D, Schumacher B (August 2015). «Genome Instability in Development and Aging: Insights from Nucleotide Excision Repair in Humans, Mice, and Worms». Biomolecules 5 (3): 1855-69. PMC 4598778. PMID 26287260. doi:10.3390/biom5031855.
- ↑ Datan E, Minn I, Peng X, He QL, Ahn H, Yu B, Pomper MG, Liu JO (2020). «A Glucose-Triptolide Conjugate Selectively Targets Cancer Cells under Hypoxia». iScience 23 (9). PMID 33083765. doi:10.1016/j.isci.2020.101536.
- ↑ Oh KS, Khan SG, Jaspers NG, Raams A, Ueda T, Lehmann A, Friedmann PS, Emmert S, Gratchev A, Lachlan K, Lucassan A, Baker CC, Kraemer KH (November 2006). «Phenotypic heterogeneity in the XPB DNA helicase gene (ERCC3): xeroderma pigmentosum without and with Cockayne syndrome». Human Mutation 27 (11): 1092-103. PMID 16947863. doi:10.1002/humu.20392.
- ↑ Le May N, Dubaele S, Proietti De Santis L, Billecocq A, Bouloy M, Egly JM (February 2004). «TFIIH transcription factor, a target for the Rift Valley hemorrhagic fever virus». Cell 116 (4): 541-50. PMID 14980221. doi:10.1016/s0092-8674(04)00132-1.
Enlaces externos
[editar]- MeSH: Transcription+Factor+TFIIH (en inglés)
 Datos: Q14908082
Datos: Q14908082