
Aphidoidea
| Áfidos | ||
|---|---|---|
| Rango temporal: (Jurásico) Cretácico – Presente | ||
 Uroleucon nigrotuberculatum | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Arthropoda | |
| Clase: | Insecta | |
| Orden: | Homoptera | |
| Suborden: | Sternorrhyncha | |
| Superfamilia: | Aphidoidea | |
| Familias | ||
| ||
Los áfidos (Aphidoidea), llamados también comúnmente pulgones, son una superfamilia de insectos fitopatógenos del suborden Sternorrhyncha. Existen alrededor de 4000 especies de áfidos en una sola familia viviente, Aphididae. Hay varias familias fósiles. Unas 250 especies son plagas para los cultivos agrícolas y forestales, así como para la jardinería.
Descripción
[editar]Poseen un tamaño que oscila entre 1-10 mm. Anatómicamente, su característica más relevante es la posesión de un estilete en su aparato bucal, estructura capaz de atravesar la epidermis de las plantas hasta llegar al floema: mediante succión, los áfidos liban la savia vegetal.[1][2]
Tienen patas finas y largas, el tarso con dos uñas. La gran mayoría tienen dos tubos, cornículos, en el quinto segmento abdominal. Segregan sustancias cerosas y compuestos defensivos.[3]
Sus hábitos alimentarios son la causa de su carácter fitopatógeno: provocan un retraso en el crecimiento de las plantas huéspedes debido a su parasitismo; además, pueden transmitir agentes fitopatógenos, es decir, actúan como vectores.[3]
Ciclo vital
[editar]
La estrategia reproductiva más simple de los áfidos es tener un solo huésped en todo el año. En este puede alternar entre generaciones con reproducción sexual y asexual (holocíclico) o en cambio tener solamente reproducción asexual o partenogenética (anholocíclico). Algunas especies tienen poblaciones holocíclicas y anholocíclicas según las circunstancias, pero no se conoce ninguna especie de áfidos que tenga solamente reproducción sexual.[5] Es posible que la alternancia de generaciones sexuales y asexuales haya evolucionado repetidamente.[6]
Sin embargo, la reproducción de los áfidos es frecuentemente más compleja e incluye migraciones entre diferentes plantas huéspedes. En aproximadamente 10 % hay alternancia entre plantas huéspedes arbóreas (huésped primario) en que los pulgones pasan el invierno y plantas herbáceas (huésped secundario), donde se reproducen en gran cantidad en el verano.[3][5] Unas pocas especies tienen una casta de soldados (polifenismo) según las condiciones ambientales y algunas controlan las proporciones de los sexos de la progenie, también según las condiciones ambientales.[7]
En una estrategia reproductiva típica, solo hay hembras en la población al comienzo del ciclo estacional (si bien en algunas especies se observan machos y hembras en esta época). Los huevos que invernaron producen solo hembras que son llamadas fundadoras; son vivíparas y partenogenéticas (no hay machos y no necesitan ser fecundadas).[8] La progenie es producida por viviparidad pseudoplacentaria, que consiste en el desarrollo de los huevos carentes de yema; los embriones se alimentan de un tejido que tiene la función de placenta. Los recién nacidos emergen de la madre inmediatamente de hacer eclosión del huevo.[9] Los huevos se producen sin que haya meiosis y son partenogenéticos[10][8] y las hijas son todas clones de la madre (telitoquia).[1][9] Los embriones se desarrollan dentro de las ovariolas de la madre, así esta da a nacer ninfas hembras del primer estadio.

Como los huevos comienzan a desarrollarse inmediatamente después de la ovulación, una hembra adulta puede llevar ninfas hembras, que a su vez ya están desarrollando embriones partenogenéticos (es decir que tales ninfas nacen embarazadas con la generación siguiente). Este fenómeno permite que los áfidos se reproduzcan a gran velocidad. Los recién nacidos son muy semejantes a los adultos a excepción del tamaño. Así, la dieta de una hembra puede afectar el tamaño y el ritmo de nacimientos de más de dos generaciones (hijas y nietas).[1][11][12]
Este proceso se repite durante el verano, produciendo múltiples generaciones que típicamente viven entre 20 y 40 días. Por ejemplo algunas especies de pulgones del repollo (como Brevicoryne brassicae) pueden producir 41 generaciones de hembras en una estación. Así, una hembra nacida en la primavera podría, en teoría, llegar a producir miles de millones de descendientes, si todos sobrevivieran.[13]

En otoño los áfidos se reproducen sexualmente y producen huevos. Los factores ambientales, tales como el cambio de fotoperíodo y la temperatura, o tal vez alimento de baja calidad, lleva a las hembras a producir hembras sexuales y machos.[10] Los machos son genéticamente iguales a sus madres, excepto que en áfidos el sistema de determinación del sexo XO hace que los machos tengan un cromosoma menos que las hembras (el cromosoma sexual).[10] Estos áfidos sexuales pueden carecer de alas y aun de piezas bucales.[3] Estos machos y hembras se aparean y las hembras depositan huevos que (a diferencia de otras generaciones) se desarrollan fuera del cuerpo de la madre. Los huevos pasan el invierno y eclosionan como hembras aladas o ápteras la primavera siguiente. Un ejemplo típico de la familia es el pulgón del rosal (Macrosiphum rosae). No obstante, en climas tropicales o en invernaderos los pulgones pueden pasar muchas generaciones sin tener individuos sexuales y reproducción por huevos.[2]
Los pulgones que se reproducen asexualmente pueden tener descendientes alados y ápteros que son genéticamente idénticos. El control es complejo; algunos áfidos alternan durante el ciclo vital entre polimorfismo (de control genético) y polifenismo (de control ambiental) en la producción de formas aladas y sin alas.[14] En general, la tendencia es de producir individuos alados cuando las condiciones son desfavorables. Algunas especies producen descendientes alados en respuesta a alimentación de baja calidad o escasa, por ejemplo cuando la planta huésped comienza a envejecer.[15] Las hembras aladas emigran para iniciar colonias en otra planta huésped. Por ejemplo, el pulgón de la manzana (Aphis pomi), después de producir muchas generaciones de hembras ápteras da lugar a formas aladas que vuelan a otras ramas u otros árboles de su planta hospedera típica.[16]
Los pulgones que son atacados por coccinélidos, neurópteros, avispas parasitoides u otros depredadores pueden cambiar la dinámica de la producción de prole. Cuando están siendo atacados producen feromonas de alarma, en particular farneseno beta, que liberan por sus cornículos. Estas feromonas inducen varios cambios de comportamiento según la especie de áfido, que pueden incluir caminar alejándose del peligro o dejarse caer de la planta. Además las feromonas de alarma pueden inducir la producción de progenie alada que puede abandonar la planta en busca de otro lugar más seguro.[17]
Las infecciones virales además de ser extremadamente dañinas para los pulgones pueden llevar a la producción de progenie alada.[18] Por ejemplo, la infección con Densovirus tiene un impacto negativo en la reproducción del pulgón de los manzanos Dysaphis plantaginea (Dysaphis plantaginea), pero contribuye al desarrollo de individuos alados, que pueden transmitir más fácilmente el virus a otras plantas huéspedes.[19] Además las bacterias simbióticas que viven en el interior de los áfidos también pueden alterar las estrategias reproductivas basadas en exposición a estreses ambientales.[20]

En otoño las especies de pulgones que tienen huéspedes alternativos (héteroecios) producen una generación alada que vuela a una planta huésped diferente donde tiene lugar la parte sexual del ciclo vital. Se producen hembras ápteras y machos y tiene lugar la producción de huevos.[21] Algunas especies, como Aphis fabae, Metopolophium dirhodum, Myzus persicae (pulgón del melocotonero) y Rhopalosiphum padi son graves plagas. Pasan el invierno en su huésped primario en árboles o arbustos; en verano emigran al huésped secundario, que generalmente es una planta herbácea, a menudo plantas de importancia agrícola; después la ginopara regresa a los árboles en otoño. Otro ejemplo es el pulgón de la soja (Aphis glycines), cuando se acerca el otoño la planta de soja envejece desde la base hacia arriba. Los pulgones emigran hacia las partes más altas de la planta y comienzan a producir generaciones aladas, primero hembras y después machos, que vuelan a su huésped primario (especies de Rhamnus) donde se aparean y producen huevos que pasan el invierno.[4]
Distribución
[editar]Los áfidos tienen una distribución mundial, pero son más comunes en las regiones templadas. En contraste con muchos otros taxones de animales, los áfidos tienen menor diversidad en los trópicos.[22] Pueden migrar grandes distancias, principalmente por dispersión pasiva por medio del viento. Los áfidos alados pueden llegar a alturas de 600 metros donde son llevados por fuertes vientos.[23][24] Por ejemplo, se cree que Nasonovia ribisnigri ha sido transportado de Nueva Zelanda a Tasmania alrededor del año 2004 por el viento.[25]
También muchos áfidos han sido transportados por los humanos con materiales vegetales, causando que algunas especies lleguen a tener distribución casi cosmopolita.[26]
7.JPG)
Evolución
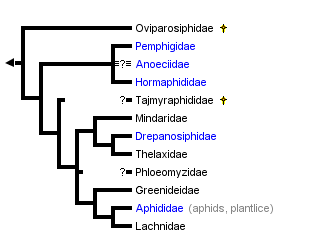
[editar]La mayoría de las familias de Aphidoidea son extintas, hay una sola familia viviente, Aphididae. Dos familias relacionadas, Phylloxeridae y Adelgidae, se encuentran situadas en la superfamilia Phylloxeroidea.
Aunque la evidencia fósil sugiere que Aphidoidea se diversificó al tiempo que lo hacían las primeras angiospermas, en el Cretácico temprano, podría ser que la familia fuera mucho más antigua. De hecho, los Prociphilini (Pemphigidae), Mindarus (Mindaridae) y Neophyllis (Drepanosiphidae) podrían ser más antiguos que las coníferas. Se considera el mínimo estimado en la edad del taxón en 140 millones de años, si bien se postula una edad de 200 millones.[27] La mayoría de las familias de áfidos se extinguieron en el límite K-T (entre el Cretácico y el Terciario), junto con los dinosaurios y otros taxones; después, radiaron en el Mioceno.[28][27]
Taxonomía
[editar]Externa
[editar]| Sternorrhyncha |
| ||||||||||||||||||||||||||||||




_(cropped).jpg)
.jpg)
Interna
[editar]| Aphididae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||

.jpg)

.jpg)
.jpg)

_on_a_rose_bud.jpg)

Nomenclatura
[editar]Existe cierta discrepancia en la nomenclatura de los taxones que subdividen al grupo. Por ejemplo, en algunos casos se prefiere una denominación de subfamilias, (con sufijo '-inae') a la de familias (sufijo '-idae'). La jerarquización taxonómica es difícil hoy día debido a la gran disparidad de enfoques y a la falta de algunos elementos clave en el registro fósil.[29]
Referencias
[editar]- ↑ a b c Dixon, A. F. G. (1998). Aphid Ecology (2nd edición). Chapman and Hall. ISBN 978-0-412-74180-7.
- ↑ a b Stroyan, Henry G. (1997). «Aphid». McGraw-Hill Encyclopedia of Science and Technology (8th edición). ISBN 978-0-07-911504-1.
- ↑ a b c d McGavin, George C. (1993). Bugs of the World. Infobase Publishing. ISBN 978-0-8160-2737-8.
- ↑ a b Wang, C. L.; Siang, L. Y.; Chang, G. S.; Chu, H. F. (1962). «Studies on the soybean aphid, Aphis glycines Matsumura». Acta Entomologica Sinica 11: 31-44.
- ↑ a b van Emden, Helmut F.; Harrington, Richard (2017). Aphids as Crop Pests, 2nd Edition. CABI. pp. 81-82. ISBN 978-1-78064-709-8.
- ↑ Von Dohlen, Carol; Moran, Nancy A. (2000). «Molecular data support a rapid radiation of aphids in the Cretaceous and multiple origins of host alternation». Biological Journal of the Linnean Society 71 (4): 689-717. doi:10.1006/bijl.2000.0470.
- ↑ Moran, Nancy A. (1992). «The Evolution of Aphid Life Cycles». Annual Review of Entomology 37: 321-348. doi:10.1146/annurev.en.37.010192.001541.
- ↑ a b Blackman, Roger L. (2008). «Stability and variation in aphid clonal lineages». Biological Journal of the Linnean Society 11 (3): 259-277. doi:10.1111/j.1095-8312.1979.tb00038.x.
- ↑ a b Gullan, P. J.; Cranston, P. S. (2010). The Insects: An Outline of Entomology (4th edición). Wiley. pp. 150–151. ISBN 978-1-118-84615-5.
- ↑ a b c Hales, Dinah F.; Wilson, Alex C. C.; Sloane, Mathew A.; Simon, Jean-Christophe; Legallic, Jean-François; Sunnucks, Paul (2002). «Lack of Detectable Genetic Recombination on the X Chromosome During the Parthenogenetic Production of Female and Male Aphids». Genetics Research 79 (3): 203-209. PMID 12220127. doi:10.1017/S0016672302005657.
- ↑ Nevo, Ettay; Coll, Moshe (2001). «Effect of nitrogen fertilization on Aphis gossypii (Homoptera: Aphididae): variation in size, color, and reproduction». Journal of Economic Entomology 94 (1): 27-32. PMID 11233124. S2CID 25758038. doi:10.1603/0022-0493-94.1.27.
- ↑ Jahn, Gary C.; Almazan, Liberty P.; Pacia, Jocelyn B. (2005). «Effect of nitrogen fertilizer on the intrinsic rate of increase of the rusty plum aphid, Hysteroneura setariae (Thomas) (Homoptera: Aphididae) on rice (Oryza sativa L.)». Environmental Entomology 34 (4): 938-943. S2CID 1941852. doi:10.1603/0046-225X-34.4.938. Archivado desde el original el 9 de septiembre de 2010.
- ↑ Hughes, R. D. (1963). «Population Dynamics of the Cabbage Aphid, Brevicoryne brassicae (L.)». Journal of Animal Ecology 32 (3): 393-424. JSTOR 2600. doi:10.2307/2600.
- ↑ Brisson, Jennifer A. (2010). «Aphid wing dimorphisms: linking environmental and genetic control of trait variation». Philosophical Transactions of the Royal Society B: Biological Sciences 365 (1540): 605-616. PMC 2817143. PMID 20083636. doi:10.1098/rstb.2009.0255.
- ↑ Weisser, Wolfgang W.; Zytynska, Sharon E.; Mehrparvar, Mohsen (5 de marzo de 2013). «Multiple Cues for Winged Morph Production in an Aphid Metacommunity». PLOS ONE (en inglés) 8 (3): e58323. Bibcode:2013PLoSO...858323M. ISSN 1932-6203. PMC 3589340. PMID 23472179. doi:10.1371/journal.pone.0058323.
- ↑ Lees, A. D. (1 de febrero de 1967). «The production of the apterous and alate forms in the aphid Megoura viciae Buckton, with special reference to the rôle of crowding». Journal of Insect Physiology 13 (2): 289-318. ISSN 0022-1910. doi:10.1016/0022-1910(67)90155-2.
- ↑ Kunert, Grit; Otto, Susanne; Röse, Ursula S. R.; Gershenzon, Jonathan; Weisser, Wolfgang W. (28 de abril de 2005). «Alarm pheromone mediates production of winged dispersal morphs in aphids». Ecology Letters 8 (6): 596-603. ISSN 1461-023X. doi:10.1111/j.1461-0248.2005.00754.x.
- ↑ Ryabov, E. V.; Keane, G.; Naish, N.; Evered, C.; Winstanley, D. (13 de mayo de 2009). «Densovirus induces winged morphs in asexual clones of the rosy apple aphid, Dysaphis plantaginea». Proceedings of the National Academy of Sciences 106 (21): 8465-8470. Bibcode:2009PNAS..106.8465R. ISSN 0027-8424. PMC 2688996. PMID 19439653. doi:10.1073/pnas.0901389106.
- ↑ Chan, C. K. (1991). Aphid-transmitted viruses and their vectors of the world. Research Branch, Agriculture Canada. ISBN 0662183347. OCLC 872604083. (requiere registro).
- ↑ Reyes, Miguel L.; Laughton, Alice M.; Parker, Benjamin J.; Wichmann, Hannah; Fan, Maretta; Sok, Daniel; Hrček, Jan; Acevedo, Tarik et al. (31 de enero de 2019). «The influence of symbiotic bacteria on reproductive strategies and wing polyphenism in pea aphids responding to stress». Journal of Animal Ecology 88 (4): 601-611. ISSN 0021-8790. PMC 6453707. PMID 30629747. doi:10.1111/1365-2656.12942.
- ↑ Alford, David V. (2014). Pests of Fruit Crops: A Colour Handbook, Second Edition. CRC Press. pp. 71-72. ISBN 978-1-4822-5421-1.
- ↑ Żyła, Dagmara; Homan, Agnieszka; Wegierek, Piotr (2017). «Polyphyly of the extinct family Oviparosiphidae and its implications for inferring aphid evolution (Hemiptera, Sternorrhyncha)». PLOS ONE 12 (4): e0174791. Bibcode:2017PLoSO..1274791Z. PMC 5405925. PMID 28445493. doi:10.1371/journal.pone.0174791.
- ↑ Berry, R. E.; Taylor, L. R. (1968). «High-Altitude Migration of Aphids in Maritime and Continental Climates». Journal of Animal Ecology 37 (3): 713-722. JSTOR 3084. doi:10.2307/3084.
- ↑ Isard, Scott A.; Irwin, Michael E.; Hollinger, Steven E. (1 de octubre de 1990). «Vertical Distribution of Aphids (Homoptera: Aphididae) in the Planetary Boundary Layer». Environmental Entomology 19 (5): 1473-1484. doi:10.1093/ee/19.5.1473.
- ↑ Hill, L. (2012). «The currant lettuce aphid, Nasonovia ribisnigri arrives in Tasmania: Part 1». Victorian Entomologist 42 (2): 29-31.
- ↑ Margaritopoulos, John T.; Kasprowicz, Louise; Malloch, Gaynor L.; Fenton, Brian (11 de mayo de 2009). «Tracking the global dispersal of a cosmopolitan insect pest, the peach potato aphid». BMC Ecology 9: 13. PMC 2687420. PMID 19432979. doi:10.1186/1472-6785-9-13.
- ↑ a b Heie, O. E. 1987. Palaeontology and phylogeny. Aphids: Their biology, natural enemies, and control, vol 2A. Eds. A. K. Minks and P. Harrewijn. Elsevier, Amsterdam.
- ↑ Johnson, Christine; Agosti, Donat; Delabie, Jocques H.; Dumpert, Klaus; Williams, D. J.; von Tschirnhaus, Michael; Macshwitz, Ulrich (2001). «Acropyga and Azteca ants (Hymenoptera: Formicidae) with scale insects (Sternorrhyncha: Coccoidea): 20 million years of intimate symbiosis». American Museum Novitates 3335: 1-18. doi:10.1206/0003-0082(2001)335<0001:AAAAHF>2.0.CO;2.
- ↑ David L. Stern y Nancy A. Moran. «Aphidoidea» (en inglés). Consultado el 28 de agosto de 2007.
Enlaces externos
[editar]- Árbol filogenético del grupo según el proyecto Tree of Life. (en inglés)
{kind=link}
| Control de autoridades |
|
|---|
 Datos: Q183350
Datos: Q183350 Multimedia: Aphidoidea / Q183350
Multimedia: Aphidoidea / Q183350 Especies: Aphidoidea
Especies: Aphidoidea