
Dictyostelea
| Dictyostelea | ||
|---|---|---|
 Moho mucilaginoso de Dictyostelium discoideum. | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| Filo: | Amoebozoa | |
| Subfilo: | Conosa | |
| Infrafilo: | Mycetozoa | |
| Clase: | Dictyostelea | |
| Orden: | Dictyostelida | |
| Familias y géneros[1] | ||
Dictyostelea o Dictyostelida es un grupo de mixomicetos, protistas conocidos comúnmente como mohos mucilaginosos.[2][3] Los dictiostélidos son amebas sociales que se alimentan de bacterias y que abundan en el suelo sobre todo cuando este es rico en material orgánico. Estos organismos pueden tomar tres formas distintas durante el transcurso de su vida: mixamebas unicelulares, una etapa de agregación denominada seudoplasmodio y un cuerpo fructífero formador de esporas.[4] Los dictiostélidos se distinguen de otros mixomicetos en que el agrupamiento se hace a partir de amebas individuales, las cuales no se fusionan, sino que cada célula mantiene su independencia al agregarse, formándose un seudoplasmodio.[5][6] El grupo comprende unas 150 especies conocidas.[7][8]
Características[editar]

Los dictiostélidos viven en los suelos del bosque, campos agrícolas, en extiercol o material en descomposión. Abundan en las zonas templadas, aunque también han sido encontrados en zonas tan frías como Alaska y en desiertos. Se alimentan por fagocitosis de bacterias, levaduras y de otros pequeños organismos. En el laboratorio pueden ser alimentados con las bacterias Escherichia coli o Klebsiella aerogenes.[9][10]
Los dictiostélidos presentan un ciclo vital que alterna entre etapas unicelulares y multicelulares. La etapa trófica (la que se alimenta) es estrictamente unicelular y consiste en mixamebas que se multiplican por fisión binaria. Sin embargo, los dictiostélidos tienen tres posibles respuestas cuando las condiciones ambientales son desfavorables o el alimento escasea. Estas son la formación de microquistes, macroquistes y de cuerpos fructíferos. El primer tipo es la formación de quistes individuales por parte de las mixamebas, de forma similar a los producidos por otros protistas.
Los macroquistes tienen dos funciones, de resistencia y como etapa sexual. Probablemente estaba presente en el antecesor de estor organismos, si bien actualmente en algunas especies no ha sido observada. Una vez formado un cigoto, este atrae a otras mixamebas circundantes que se fusionan para formar pequeños agregados. Los cigotos aumentan progresivamente de tamaño y se rodean de una cubierta que protege a todo el colectivo. Antes de la germinación, la célula gigante se divide para volver a dar lugar a mixamebas uninucleadas.
Sin embargo, la etapa más notable es la formación de cuerpos fructíferos. Esta es una verdadera etapa multicelular consistente en la agregación de numerosas mixamebas para formar un seudoplasmodio capaz de desplazarse sobre el sustrato y eventualmente da lugar a un cuerpo fructífero, compuesto de un tallo de celulosa que soporta un esporocarpo. Sorprendentemente, los dictióstelidos han evolucionado para producir varios tipos de células diferenciadas, un proceso que incluye la capacidad de regular las proporciones y morfogénesis de cada tipo.
Algunas especies presentan fototaxis, de forma que los cuerpos fructíferos se localizan en los lugares más adecuados para dispersar las esporas. Esta se realiza principalmente a través de pequeños animales como insectos, anfibios, roedores, aves y murciélagas.
-
 Dictyostelium, 1-mixamebas
Dictyostelium, 1-mixamebas -
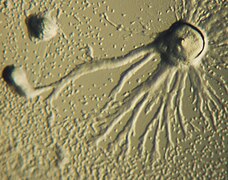 Dictyostelium, 2-comienzo de la agregación
Dictyostelium, 2-comienzo de la agregación -
 Dictyostelium, 3-final de la agregación
Dictyostelium, 3-final de la agregación -
 Dictyostelium, 4-seudoplasmodio
Dictyostelium, 4-seudoplasmodio -
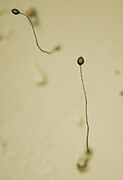 Dictyostelium, 5-cuerpos fructíferos
Dictyostelium, 5-cuerpos fructíferos





Ciclo de vida[editar]

El ciclo de vida de estos organismos comprende las etapas de espora, mixameba, seudoplasmodio y cuerpo fructífero.
La espora es la fase de reposo y de dispersión de estos organismos. Las esporas son células inactivas protegidas por paredes celulares resistentes.
De las esporas se liberan mixamebas unicelulares y microscópicas, de unas 5 µm de longitud, que se alimentan de bacterias y se dividen normalmente por mitosis.
Sin embargo, cuando la fuente de alimento se agota, las mixamebas se agrupan para formar un agregado multicelular denominado seudoplasmodio. El seudoplasmodio puede desplazarse con partes anterior y posterior definidas, respondiendo a la luz y a los gradientes de temperatura. Normalmente la agrupación consta de 10.000 a 100.000 individuos, aunque se han observado agrupaciones que van desde 100 células hasta 2.000.000. El tamaño del seudoplasmodio suele estar comprendido entre 0,5 y 2,0 mm.
Bajo condiciones apropiadas, los seudoplasmodios maduros forman un cuerpo fructífero portando una o más bolas de esporas soportadas por los correspondientes tallos. En Acytostelium, el cuerpo fructífero está soportado por un tallo compuesto de celulosa, pero en otros dictiostélidos el tallo está formado por células, que, excepto en unos pocos casos, mueren durante la formación del tallo. Hay una correspondencia definida entre las partes del seudoplasmodio y las del cuerpo fructífero. Las esporas se transforman en nuevas amebas cuando el alimento vuelve a estar disponible.
Mecanismo de agregación[editar]

La agregación de las amebas generalmente tiene lugar en la forma de flujos convergentes. Las amebas se mueven usando seudópodos filosos y son atraídas por los compuestos químicos producidos por otras amebas. En Dictyostelium, el mecanismo que permite la agregación de las amebas se basa en el adenosín monofosfato cíclico (AMPc) como molécula señalizadora, pero otras especies usan diferentes compuestos químicos. En la especie Dictyostelium purpureum, el agrupamiento implica también parentesco, no solo por proximidad.
El preceso de agregación tiene lugar de la siguiente forma. Una célula, la fundadora de la colonia, comienza a segregar AMPc en respuesta al estrés. Las demás detectan esta señal y responden realizando dos acciones:
- La ameba se mueve hacia la señal.
- La ameba segrega más AMPc para reforzar la señal.
El efecto de esto es la retransmisión de la señal a través de la población de amebas cercanas y las dirige hacia el área de mayor concentración de AMPc. Con más detalle y para cada célula individual, el mecanismo funciona como sigue:
- La recepción de AMPc en la membrana celular activa una proteína G.
- La proteína G estimula la adenilato ciclasa.
- Se difunde AMPc desde la célula al medio.
- El AMPc interno inactiva el receptor de AMPc externo.
- Una proteína G distinta estimula la fosfolipasa C.
- IP3 induce la segregación de iones de calcio.
- Los iones de calcio actúan sobre el citoesqueleto para inducir la extensión de seudópodos.
Debido a que la concentración interna de AMPc inactiva el receptor de AMPc externo, una célula individual presenta un comportamiento oscilatorio. Este comportamiento produce las bonitas espirales que se ven en las colonias convergiendo, lo que es un reminiscencia de la reacción Belousov-Zhabotinsky y de los autómatas celulares cíclicos.
Genoma[editar]
El genoma completo de Dictyostelium contiene aproximadamente 12.500 genes en seis cromosomas. Por comparación, el genoma humano contiene 20.000-25.000 genes en 23 pares de cromosomas. El genoma contiene una alta proporción de los nucleótidos adenosina y timidina (~77%) lo que favorece el uso de codones que contienen en su tercera posición adenosina y timina. En Dictyostelium, repeticiones de trinucleótidos son también abundantes, mientras que en los seres humanos esta repetición de trinucleótidos ocasiona enfermedades.
Utilidad en investigación[editar]
Dictyostelium se ha utilizado como organismo modelo en biología molecular y genética, y ha sido estudiado como ejemplo de comunicación celular, diferenciación y muerte programada. Es también un interesante ejemplo de la evolución de las cooperación y de las estrategias tramposas.[11][12][13]
Referencias[editar]
- ↑ Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
- ↑ Fiore-Donno, A. M., Nikolaev, S. I., Nelson, M., Pawlowski, J., Cavalier-Smith, T., & Baldauf, S. L. (2010). Deep phylogeny and evolution of slime moulds (Mycetozoa). Protist, 161(1), 55-70.
- ↑ Baldauf, S. L., & Doolittle, W. F. (1997). Origin and evolution of the slime molds (Mycetozoa). Proceedings of the National Academy of Sciences, 94(22), 12007-12012.
- ↑ Schnittler, M., Novozhilov, Y. K., Romeralo, M., Brown, M., & Spiegel, F. W. (2012). Fruit body-forming protists: Myxomycetes and myxomycete-like organisms. Englers Syllabus of Plant Families, 1, 1.
- ↑ Baldauf SL, Doolittle WF (October 1997). «Origin and evolution of the slime molds (Mycetozoa)». Proc. Natl. Acad. Sci. U.S.A. 94 (22): 12007-12. PMC 23686. PMID 9342353. doi:10.1073/pnas.94.22.12007.
- ↑ Schnittler, M., Novozhilov, Y. K., Romeralo, M., Brown, M., & Spiegel, F. W. (2012). Fruit body-forming protists: Myxomycetes and myxomycete-like organisms. Englers Syllabus of Plant Families, 1, 1.
- ↑ Ald, S.M. et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ↑ Chapman, A. D. (2009). Numbers of living species in Australia and the world Archivado el 28 de septiembre de 2015 en Wayback Machine..
- ↑ Romeralo, M., Escalante, R., & Baldauf, S. L. (2012). Evolution and diversity of dictyostelid social amoebae. Protist, 163(3), 327-343.
- ↑ Romeralo, M., Fiz-Palacios, O., Lado, C., & Cavender, J. C. (2007). A new concept for Dictyostelium sphaerocephalum based on morphology and phylogenetic analysis of nuclear ribosomal internal transcribed spacer region sequences. Botany, 85(1), 104-110.
- ↑ Strassman JE, Zhu Y, and Queller DC. (2000) Altruism and social cheating in the social amoeba Dictyostelium discoideum. Nature.
- ↑ Dao DN, Kessin RH, and Ennis HL (2000). Developmental cheating and the evolutionary biology of Dictyostelium and Myxococcus. Microbiology.
- ↑ Brännsröm Å and Dieckmann U (2005). Evolutionary dynamics of altruism and cheating among social amoebas. Proceedings of the Royal Society of London B.
Véase también[editar]
Enlaces externos[editar]
 Wikispecies tiene un artículo sobre Dictyostelea.
Wikispecies tiene un artículo sobre Dictyostelea. Wikimedia Commons alberga una categoría multimedia sobre Dictyosteliida.
Wikimedia Commons alberga una categoría multimedia sobre Dictyosteliida.- The genome of the social amoeba Dictyostelium discoideum (2005) Nature 435, 43-57
- Dictyostelium (2007)
- Low Society (2004)
- DictyBase y DictyBase wiki (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).. Online Informatics Resource for Dictyostelium.
- Dictyostelium discoideum Genome Project Archivado el 6 de diciembre de 2019 en Wayback Machine.
- Dictyostelium discoideum: una amiba para todas y todas para una en Revista Digital La Hoja de Arena
- Protocols for growing Dictyostelium discoideum. Cómo cultivar Dictyostelium (Consultado el 1/1/2016).
| Control de autoridades |
|
|---|
 Datos: Q310364
Datos: Q310364- Multimedia: Dictyosteliida / Q310364
- Especies: Dictyosteliida