
Células T auxiliares foliculares B
Las células T auxiliares foliculares B (también conocidas como células T auxiliares foliculares o TFH), son células T CD4+ experimentadas con antígeno que se encuentran en la periferia dentro de los folículos de células B de órganos linfoides secundarios, como ganglios linfáticos, bazos y placas de Peyer, y se identifican por su expresión constitutiva del receptor de folículo de células B CXCR5. Tras la interacción celular y la señalización cruzada con sus células B foliculares (Fo B) afines, las células TFH desencadenan la formación y el mantenimiento de los centros germinales a través de la expresión del ligando CD40 (CD40L) y la secreción de IL-21[1] e IL-4. Las células TFH también migran hacia estos centros germinales sembrados, compuestos predominantemente por células B que se dividen y mutan rápidamente. Dentro de los centros germinales, las células TFH desempeñan un papel fundamental en la mediación de la selección y la supervivencia de las células B que se diferencian en células plasmáticas especiales capaces de producir anticuerpos de alta afinidad contra antígenos extraños, o células B de memoria capaces de una rápida respuesta inmunológica en el futuro si alguna vez se vuelve a encontrar el mismo antígeno. También se cree que las células TFH facilitan la selección negativa de células B mutadas que pueden ser autoinmunes en el centro germinal. Sin embargo, los biomecanismos por los cuales las células TFH median la tolerancia del centro germinal aún no se han entendido completamente.
Es posible que las células TFH puedan surgir como ramas en las vías de diferenciación Th1 y Th2, pero su relación de linaje precisa con los otros subconjuntos de células T CD4+ efectoras aún es incierta. Sin embargo, estudios recientes han demostrado que TFH tiene distintos perfiles de expresión génica, lo que respalda la teoría de que T FH es un subconjunto de células T CD4+ distintas de Th-1, Th-2, Th-17 o Tregs.[2][3]
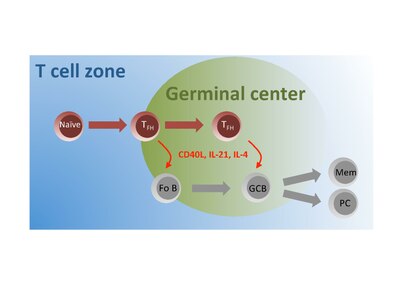
Un subconjunto de células T inmaduras en la zona de células T son activadas por el antígeno y migran a los folículos donde se diferencian en células TFH que interactúan e instruyen a las células B foliculares (Fo B) para que experimenten un cambio de isotipo, hipermutación somática y células rápidas. división a los centros germinales de semillas (GC). Dentro de estos centros germinales, las células T FH continúan brindando ayuda a las células GC B para facilitar su producción de células plasmáticas (PC) productoras de anticuerpos de alta afinidad y células B de memoria (Mem) de larga vida.
Caracterización biomolecular[editar]
Se ha demostrado que el coestimulador inducible de células T (CD278 o ICOS) proporciona una señal particularmente crítica para las células TFH ya que los ratones experimentales deficientes en ICOS no pueden desarrollar ninguna TFH.[4] Además, se ha demostrado que ICOS induce la secreción de citocina IL-21 por las células T CD4+ activadas y que IL-21 juega un papel crucial en el desarrollo de células TFH y centros germinales.[5][6] También Bcl-6 es un factor de transcripción identificado en las células TFH, pero puede tener funciones que se extienden más allá de este subconjunto, porque también se ha implicado en el desarrollo de células T CD8+ de memoria.[7]
En los centros germinales, las células TFH con antígeno regulan rápidamente la expresión de CD40L, que se une y estimula el receptor de superficie de la célula B CD40.[8] La activación paracrina dependiente de células TFH de CD40 de células B da como resultado la supervivencia y diferenciación de células B, incluida la inducción de AID ( desaminasa inducida por activación (citidina)).[9] La expresión de AID (codificada por el gen AICDA) hace que los anticuerpos de células B cambien de clase de IgM/IgD a otros isotipos de anticuerpos e impulsa la hipermutación somática durante la proliferación clonal. Los anticuerpos conmutados adquieren mejores funciones efectoras, y el anticuerpo hipermutado muestra una mayor afinidad por el antígeno.
Clases de células TFH[editar]
Las células TFH formadas temprano en las etapas nacientes de una reacción del centro germinal se denominan formalmente células pre-TFH. Se encuentran principalmente en el borde de la zona de células T que se fusiona con los folículos de células B y los centros germinales. Las células Pre-TFH son funcionalmente muy similares a otras células TFH para facilitar las reacciones de las células B del centro germinal, sin embargo, además, también son capaces de impulsar el desarrollo de células B foliculares adyacentes y fuera de los centros germinales para producir una respuesta rápida pero no respuestas duraderas de anticuerpos dirigidos por células plasmáticas (conocidas como la respuesta extrafolicular).
Esas células TFH que residen específicamente dentro de un centro germinal maduro a veces se denominan células GC TFH (para las células TFH del centro germinal) para distinguirlas de las células pre-TFH.[10][11] También hay una subclase menor dentro de esta población de células GC Tfh que expresan el gen Foxp3, que codifica un factor de transcripción. Esta pequeña subpoblación discreta de células, llamadas células TFR (para células reguladoras foliculares T), es importante para ayudar a controlar y limitar la magnitud de las respuestas normales del centro germinal de modo que eviten la posibilidad de producir mutaciones anormales o autorreactivas anticuerpos autoinmunes asociados.[12] Por lo tanto, las células TFR son una influencia inhibitoria única durante una reacción del centro germinal.
Mientras que las células TFH se encuentran principalmente en los órganos linfoides secundarios, una pequeña proporción circula en la sangre y se denominan células auxiliares foliculares T "periféricas" (pTFH). Estas células pueden identificarse por su expresión de IL-21 tras la estimulación.[13]
Relevancia médica[editar]
Generando memoria inmune duradera[editar]
Las células TFH se consideran un subconjunto indispensable de células T en la generación y mantenimiento de las respuestas del centro germinal. Por lo tanto, en ausencia de células TFH, similar a la activación de células B por antígenos independientes de células T, se forma una explosión rápida de producción de células plasmáticas de baja afinidad, pero esto no conduce a la inducción del centro germinal ni permite la maduración por afinidad de anticuerpos o la diferenciación de células B de memoria eficaces que son esenciales para fortalecer el cuerpo contra infecciones posteriores. Específicamente, las células B de memoria dependientes del centro germinal son los impulsores de la producción de anticuerpos de recuerdo durante una respuesta inmune secundaria. Por lo tanto, la activación y el desarrollo adecuados de las células TH son fundamentales para la eficacia de las inmunizaciones y el diseño de vacunas para la inducción de inmunidad a largo plazo. En un estudio de la población de Bangladés de pacientes infectados con Vibrio cholera y voluntarios humanos sanos administrados con una vacuna contra el cólera existente,[14] una respuesta de memoria TFH específicamente contra el antígeno del cólera se había correlacionado con una mayor producción de anticuerpos por parte de las células B.
Controlar el deterioro inmunitario relacionado con la edad[editar]
Con el envejecimiento normal viene una disminución gradual del sistema inmune del cuerpo. Este fenómeno llamado inmunosenescencia se debe en gran medida a una disminución de la función de las células T, incluida la capacidad de las células TFH para soportar adecuadamente las respuestas del centro germinal.[15] Esto puede deberse en parte a los niveles más bajos de CD40L en la superficie celular de las células TFH en los ancianos.[16]
Evitar la autoinmunidad[editar]
Las respuestas inmunitarias no controladas o hiperactivas de las células TFH tienen el potencial de montar centros germinales injustificados, compuestos de células B mutadas aberrantemente que pueden conducir enfermedades autoinmunes mediadas por anticuerpos. Se pueden detectar niveles elevados de células similares a TFH en la sangre de un subconjunto de pacientes humanos con lupus eritematoso sistémico (LES) y síndrome de Sjogren.[17] Sin embargo, la evidencia científica que sugiere que las células TFH pueden causar definitivamente autoinmunidad en humanos permanece incompleta.
Referencias[editar]
- ↑ Seo; Youn, J; Kim, PH (May 2009). «IL-21 ensures TGF-beta 1-induced IgA isotype expression in mouse Peyer's patches». J Leukoc Biol 85 (5): 744-50. PMID 19168593. doi:10.1189/jlb.0708450.
- ↑ Chtanova; Tangye, SG; Newton, R; Frank, N; Hodge, MR; Rolph, MS; MacKay, CR (July 2004). «T follicular helper cells express a distinctive transcriptional profile, reflecting their role as non-Th1/Th2 effector cells that provide help for B cells». J Immunol 173 (1): 68-78. PMID 15210760. doi:10.4049/jimmunol.173.1.68.
- ↑ «Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages». Immunity 29 (1): 138-49. 2008. PMC 2556461. PMID 18599325. doi:10.1016/j.immuni.2008.05.009.
- ↑ Akiba; Takeda, K; Kojima, Y; Usui, Y; Harada, N; Yamazaki, T; Ma, J; Tezuka, K et al. (Aug 2005). «The role of ICOS in the CXCR5+ follicular B helper T cell maintenance in vivo». J Immunol 175 (4): 2340-8. PMID 16081804. doi:10.4049/jimmunol.175.4.2340.
- ↑ Bauquet; Jin, H; Paterson, AM; Mitsdoerffer, M; Ho, IC; Sharpe, AH; Kuchroo, VK (Feb 2009). «The costimulatory molecule ICOS regulates the expression of c-Maf and IL-21 in the development of follicular T helper cells and TH-17 cells». Nat Immunol 10 (2): 167-75. PMC 2742982. PMID 19098919. doi:10.1038/ni.1690.
- ↑ Vogelzang; McGuire, HM; Yu, D; Sprent, J; MacKay, CR; King, C (July 2008). «A fundamental role for interleukin-21 in the generation of T follicular helper cells». Immunity 29 (1): 127-37. PMID 18602282. doi:10.1016/j.immuni.2008.06.001.
- ↑ Ichii; Sakamoto, A; Hatano, M; Okada, S; Toyama, H; Taki, S; Arima, M; Kuroda, Y et al. (June 2002). «Role for Bcl-6 in the generation and maintenance of memory CD8+ T cells». Nat Immunol 3 (6): 558-63. PMID 12021781. doi:10.1038/ni802.
- ↑ Kim, JR; Lim, HW; Kang, SG; Hillsamer, P; Kim, CH (4 de febrero de 2005). «Human CD57+ germinal center-T cells are the major helpers for GC-B cells and induce class switch recombination.». BMC Immunology 6: 3. PMC 548684. PMID 15694005. doi:10.1186/1471-2172-6-3.
- ↑ McHeyzer-Williams, M; Okitsu, S; Wang, N; McHeyzer-Williams, L (Dec 9, 2011). «Molecular programming of B cell memory.». Nature Reviews. Immunology 12 (1): 24-34. PMC 3947622. PMID 22158414. doi:10.1038/nri3128.
- ↑ «Germinal center T follicular helper cell IL-4 production is dependent on signaling lymphocytic activation molecule receptor (CD150).». J. Immunol. 185 (1): 190-202. July 2010. PMC 2913439. PMID 20525889. doi:10.4049/jimmunol.0903505.
- ↑ «T-cell subsets in the germinal center.». Immunol. Rev. 252 (1): 146-55. March 2013. PMID 23405902. doi:10.1111/imr.12031.
- ↑ «Regulation of the germinal center reaction by Foxp3+ follicular regulatory T cells.». J. Immunol. 187 (9): 4553-60. November 2011. PMID 21984700. doi:10.4049/jimmunol.1101328.
- ↑ Schultz, Bruce T.; Teigler, Jeffrey E.; Pissani, Franco; Oster, Alexander F.; Kranias, Gregory; Alter, Galit; Marovich, Mary; Eller, Michael A. et al. (19 de enero de 2016). «Circulating HIV-Specific Interleukin-21(+)CD4(+) T Cells Represent Peripheral Tfh Cells with Antigen-Dependent Helper Functions». Immunity 44 (1): 167-178. ISSN 1097-4180. PMID 26795249. doi:10.1016/j.immuni.2015.12.011.
- ↑ «Antigen-specific memory T cell responses after vaccination with an oral killed cholera vaccine in Bangali children and comparison to responses in patients with naturally acquired cholera.». Clinical and Vaccine Immunology 19 (8): 1304-11. August 2012. PMC 3416086. PMID 22739692. doi:10.1128/CVI.00196-12.
- ↑ «The aged microenvironment contributes to the age-related functional defects of CD4 T cells in mice.». Aging Cell 11 (5): 732-40. 2012. PMC 3444657. PMID 22607653. doi:10.1111/j.1474-9726.2012.00836.x.
- ↑ «Ex vivo enzymatic treatment of aged CD4 T cells restores cognate T cell helper function and enhances antibody production in mice.». J Immunol 189 (12): 5582-9. 2012. PMC 3518580. PMID 23136198. doi:10.4049/jimmunol.1200487.
- ↑ Craft JE. (2012). «Follicular helper T cells in immunity and systemic autoimmunity.». Nat Rev Rheumatol 8 (6): 337-47. PMC 3604997. PMID 22549246. doi:10.1038/nrrheum.2012.58.
| Control de autoridades |
|
|---|
 Datos: Q5464650
Datos: Q5464650