
ARN polimerasa V
La ARN polimerasa V, a veces abreviada como Pol V, es un tipo específico de ARN polimerasa que se encuentra únicamente en el reino vegetal. En todas las células el ADN nuclear aporta información para la codificación del ARN, pero para llevar a cabo la síntesis es imprescindible también la actuación de las ARN polimerasas. En las células procariotas un mismo tipo de ARN polimerasa cataliza la síntesis de todos los tipos de ARN (ARNm, ARNt y ARNn). Las células eucariotas, en cambio, han desarrollado diferentes tipos de ARN polimerasa, cada uno especializado en funciones diferentes. Los más conocidos y estudiados son el ARN polimerasa I, II y III, especialmente el II. Sin embargo, también existen, aunque menos estudiados, dos tipos más de ARN polimerasa que se encuentran exclusivamente en las plantas: el ARN polimerasa IV y el V.
Estos dirigen el ARN no codificante en la regulación de algunos procesos relacionados con el silenciamiento de algunos genes implicados en el desarrollo, el control de la movilidad de los transposones, la defensa contra los virus y el cruzamiento entre alelos. Aún son desconocidos los detalles de la naturaleza bioquímica de estos dos tipos de ARN polimerasa, pero lo poco que se conoce de sus composiciones revela que podrían haberse desarrollado a partir del ARN polimerasa II como una forma especializada de este tipo.
El interés en el estudio del ARN polimerasa IV y V radica en la información que aporta sobre la diversificación funcional en una misma familia de enzimas.
Polimerasa V y Arabidopsis[editar]
Estudios hechos en plantas del género de los Arabidopsis (género de plantas herbáceas de la familia de las brasicáceas) muestran que la Pol V está relacionada con la organización de la heterocromatina y el silenciamiento de genes que son independientes del 24-nt siRNA (dependiente de polimerasa IV) y del DRM2 (dependiente de metilación). En la metilación, la Pol V y DRD1 - un remodelador de cromatina requerido para la transcripción de la Pol V - aparentemente actúan en compañía de MET1 y DDM1, la actividad de los cuales es necesaria para mantener la mayoría de la metilación en el genoma de los Arabidopsis. Este proceso para generar metilación no es muy bien conocido todavía, pero la última hipótesis para explicar este suceso consiste en la generación de transcripciones en el locus de la heterocromatina que provoca la unión a los complejos siRNA-AGO4. Como consecuencia de esta unión, se produce el reclutamiento de metilación del DNA y de maquinaria modificadora de histonas. Por otro lado, en plantas de la familia de los Arabidopsis también observamos que la Pol V, independientemente de la Pol IV, RDR y la producción de siRNA mediante DCL3, afecta a la organización de la heterocromatina durante la interfase, posiblemente envolviendo los ARN de proteínas estructurales o modificadoras de la cromatina. Cabe destacar la técnica ‘’in vivo’’ de immunoprecipitación como una de las que más información aportaron sobre la función de la Pol V.
Silenciación de genes[editar]
La secuenciación del genoma de Arabidopsis thaliana dio lugar al descubrimiento de dos nuevas secuencias de las primeras dos subunidades más grandes que fomarían parte de dos ARN polimerasas potenciales distintas de las anteriormente conocidas polimerasas I, II y III. El hecho de que estas dos subunidades formaran el centro activo de las para entonces conocidas polimerasas dio pie a la sugestión de la existencia de dos formas atípicas de polimerasa. Análisis filogenéticos de estas concluyeron que sólo están presentes en las plantas, sugiriendo que llevarían a cabo funciones específicas de las plantas. Tras la formulación de estas afirmaciones, sus funciones se fueron definiendo como dos actividades nucleares no esenciales para la viabilidad, pero necesarias para la silenciación de genes no transcripcional mediada por ARN y la formación de heterocromatina.
La polimerasa V y su transcripción son necesarias pero no suficientes en la silenciación de genes llevada a cabo mediante un proceso de metilación del ADN dirigido por el ARN. Otras proteínas tales como la polimerasa IV, RDR2 y IDN2 también toman parte en el proceso en el que se da por un lado la síntesis de siRNA y por otro la transcripción mediada por la polimerasa V.
La identificación de ARNs no codificantes dependientes de Pol V producidos en múltiples loci sujetos a la metilación del ADN dirigido por el ARN supuso un gran avance en la comprensión del papel que juega la Pol V en este proceso. Estos ARN dependientes de Pol V abarcan las regiones promotoras de los loci silenciadores, son dependientes del centro activo de la Pol V y pueden ser químicamente reticulados a Pol V, indicando que son transcripciones directas de ella. Estas transcripciones se generan independientemente de las siRNA, por lo que la Pol IV y la V trabajan independientemente siguiendo caminos paralelos que convergen para que ocurra la metilación del ADN dirigida por el ARN.
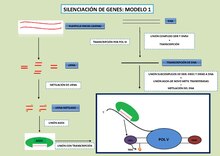
En los Arabidopsis, este proceso incluye 24 nucleótidos siRNAs que se unen a proteínas de la familia ARGONAUTE (AGO4) y facilitan el reconocimiento del loci complementario a través de mecanismos desconocidos. El ARN Polimerasa V nuclear es una enzima ARN silenciosa que genera transcripciones no codificantes en los loci silenciados por 24nt siRNAs. AGO4 interacciona con las transcripciones de Pol V y este es guiado a lugares de destino loci a través del apareamiento de bases de siRNAs asociadas a transcripciones nacientes de Pol V. AGO4 y Pol V están asociados a un cuerpo de Cajal que está alejado del loci de reconocimiento sometido a mediar el siRNA. Aún no se conoce bien la funcionalidad de esta localización en el cuerpo de Cajal, pero podría reflejar el procesamiento o complejidad de funciones independientes que inducen al complejo silenciador.
Hay diferentes hipótesis acerca del modelo de acción que seguiría la Pol V en la silenciación de genes:
- Modelo 1: Es el más aceptado y presente en la Arabidopsis thaliana. Se dan dos procesos independientes en los cuales participan la Pol IV y la Pol V respectivamente. Por un lado, la Pol IV inicia el proceso de creación de los siRNA, los cuales una vez metilados se unen con el complejo ARGONAUTE4 (AGO4). Independientemente de este proceso, la transcripción de Pol V es llevada a cabo por el complejo DDR y DMS4. Posteriormente, el complejo AGO4 unido a las siRNA interacciona con las transcripciones nacientes de Pol V dando lugar al emparejamiento de bases entre el siRNA y la transcripción. Proteínas del complejo DDR (DRD1 y DRM2) unen a AGO4 con las de novo metil transferasas acercándolas a las regiones transcritas Pol V y resultando en la metilación del ADN, proceso por el cual se produce el silenciamiento de genes.
- Modelo 2: AGO4 interacciona con la transcripción naciente, pero las bases de los siRNA se emparejan con el ADN.
- Modelo 3: el siRNA asociado al AGO4 interactúa con el ADN de manera dependiente de la perturbación en la cromatina mediada por la Pol V.
Referencias[editar]
- Pontes, Olga; Costa-Nunes, Pedro; Vithayathil, Paul; Pikaard, Craig S. (2009). «RNA Polymerase V Functions in Arabidopsis Interphase Heterochromatin Organization Independently of the 24-nt siRNA-Directed DNA Methylation Pathway». Molecular Plant 2 (4): 700–710. doi:10.1093/mp/ssp006. (enlace roto disponible en Internet Archive; véase el historial, la primera versión y la última).
- Wierzbicki, Andrzej T.; Ream, Thomas; Haag, Jeremy R.; Pikaard, Craig S. (2009). «RNA Polymerase V transcription guides ARGONAUTE4 to chromatin». Nature Genetics 41 (5): 630–634. doi:10.1038/ng.365.
- Wierzbicki, Andrzej T.; Haag, Jeremy R.; Pikaard, Craig S. (2008). «Noncoding transcription by RNA Polymerase Pol IVb/Pol V mediates transcriptional silencing of overlapping and adjacent genes». Cell. 135 (4). doi:10.1016/j.cell.2008.09.035.
- Haag, Jeremy R.; Pikaard, Craig S. (2011). «Multisubunit RNA polymerases IV and V: purveyors of non-coding RNA for plant gene silencing». Nature Reviews Molecular Cell Biology 12: 483-492. doi:10.1038/nrm3152.
Enlaces externos[editar]
| Control de autoridades |
|
|---|
 Datos: Q2886927
Datos: Q2886927