
Diferencia entre revisiones de «Ranunculales»
Sin resumen de edición |
|||
| Línea 1: | Línea 1: | ||
{{Artículo bueno}} |
|||
{{Cladobox |
{{Cladobox |
||
| color = lightgreen |
| color = lightgreen |
||
Revisión del 00:14 15 ago 2007
Ranunculales es un taxón de eudicotiledóneas ubicado en la categoría de orden, que en los sistemas de clasificación actuales como el APG (1998 [1] ), su sucesor APG II (2003 [2] ), y el más actualizado APW [3] ) está compuesto por 7 familias cuyas relaciones filogenéticas se conocen bien desde hace un tiempo. Este clado es una rama basal de las eudicotiledóneas y contiene cerca de un 1,6% de la diversidad de eudicotiledóneas.
Caracteres
Los miembros de este clado comparten los siguientes caracteres: en algunos miembros la presencia de O-metilflavonoles, presencia de flavonoles; en el corte transversal del tallo joven ("estela") se ven los haces vasculares separados, los vasos presentes sólo en la parte central, presencia de traqueidas verdaderas; los rayos exclusivamente anchamente multiseriados (y también en el leño, cuando existe); madera fluorescente en algunos representantes; cambium estratificado; las células cribosas del floema con plástidos largamente de tipo S (S-type), presencia de proteína P dispersiva (dispersive P-protein); en el corte transversal ("estela") del pecíolo los haces vasculares se disponen en forma de anillo; las ceras de la cutícula de la hoja formando grupos de túbulos, siendo la cera principal el nonacosan-10-ol; hojas dispuestas en espiral; las piezas del gineceo opuestas a las del perianto; el perianto deciduo cuando se forma el fruto; semilla con exotesta.
La berberina, de presencia común en este orden, es sintetizada por la vía de la tirosina.
A la fecha de edición de este artículo (abril del 2007), no había información suficiente sobre los elementos de vaso, los dientes de las hojas, el desarrollo del endosperma, ni el tamaño del embrión.
Filogenia
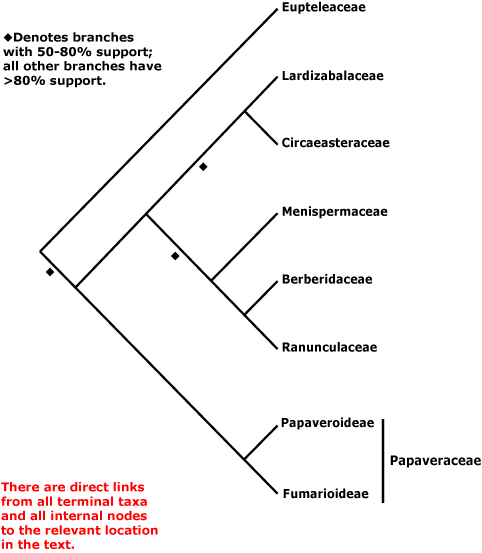
Ranunculales es el clado basal de eudicotiledóneas. Se hipotetiza que es monofilético en base al las secuencias de ADN rbcL, atpB y ADNr 18S (Chase et al. 1993[4], Drinnan et al. 1994[5], Hoot y Crane 1995[6], Loconte et al. 1995[7], Soltis et al 1997[8], 1998[9], 2000[10] , Källersjö et al. 1998[11], Hoot et al. 1999[12], Savolainen et al 2000a[13], b[14]). La presencia del alcaloide berberina también puede ser una sinapomorfía del grupo. El orden está compuesto por 7 clados a los que les fue asignada la categoría de familia, las familias más numerosas son Menispermaceae, Berberidaceae, Ranunculaceae y Papaveraceae. Estas familias tradicionalmente fueron asociadas por su hábito predominantemente herbáceo, sus hojas dentadas o lobadas o hasta compuestas, la presencia de alcaloides, típicamente del tipo de la bencil-isoquinolina, las flores hipóginas con las partes usualmente diferenciadas y libres, y usualmente con muchos estambres, y semillas con embriones pequeños y endosperma copioso (Cronquist 1981[15], Thorne 1974[16], 1992[17]).
El grupo fue usualmente asociado con el complejo de magnólidas (por ejemplo con las Magnoliales, las Laurales y las Canellales: Cronquist 1968[18], 1981[15], 1988[19], Dahlgren 1983[20],Takhtajan 1969[21], 1980[22], Thorne 1974[16], 1992[17]), pero la morfología (Donoghue y Doyle 1989[23], Doyle et al. 1994[24]) y las secuencias de ADN rbcL, atpB y ADNr 18S ya citadas indican que Ranunculales es el clado hermano de todo el resto de las tricolpadas (eudicotiledóneas).
El árbol filogenético actualizado hasta abril del 2007[25]:
| Ranunculales |
| ||||||||||||||||||||||||||||||||||||||||||||||||
|
El resto de las eudicotiledóneas | |||||||||||||||||||||||||||||||||||||||||||||||||
Papaveraceae está formada por Papaveroideae, Fumarioideae y Pteridophilloideae. Difiere del resto de los miembros del grupo en su gineceo sincarpo, frutos capsulares, sépalos tempranamente deciduos, y presencia de o bien laticíferos y savia coloreada, o bien células especializadas con savia mucilaginosa y clara. Estos caracteres, junto con las semillas con arilo, son probablemente sinapomorfías de Papaveraceae, o evolucionaron muy tempranamente en esta familia.
Las sinapomorfías morfológicas del clado que reúne a Menispermaceae, Berberidaceae y Ranunculaceae no son evidentes, aunque el grupo posee un robusto soporte por parte de las secuencias de ADN. Puede que Berberidaceae y Ranunculaceae tengan algunas similitudes en la forma de la flor (Endress 1995[26]).
Taxonomía
El orden contiene 7 familias, 199 géneros, y 4.445 species.
- Familia Ranunculaceae
- Familia Berberidaceae
- Familia Menispermaceae
- Familia Lardizabalaceae
- Familia Circaeasteraceae
- Familia Eupteleaceae
- Familia Papaveraceae (incluyendo a Fumariaceae y Pteridophyllaceae)
Existen otras pocas familias separadas del orden Ranunculaceae. En el sistema de clasificación de Cronquist (Cronquist 1981[15], 1988[19]), las Eupteleaceae eran excluidas de este orden, como también las Papaveraceae y Fumariaceae (incluyendo a las Pteridophyllaceae) que eran consideradas como parte del orden Papaverales.
Sinónimos:
- Berberidales Dumortier,
- Eupteleales Reveal,
- Glaucidiales Reveal,
- Helleborales Nakai,
- Hydrastidales Takhtajan,
- Lardizabalales Loconte,
- Menispermales Bromhead,
- Nandinales Doweld,
- Papaverales Dumortier,
- Podophyllales Dumortier - Berberidanae Doweld,
- Ranunculanae Reveal - Ranunculidae Reveal - Berberidopsida Brogniart,
- Papaveropsida Brongniart,
- Ranunculopsida Brongniart
Evolución
Magallön et al. (1999 [27]) sugieren que el clado tiene una edad de 70 Ma, basados en los fósiles más antiguos que pueden ser asignados al mismo, pero esos fósiles son miembros de Menispermaceae que no es un lineaje basal. Anderson et al. (2005[28]) datan al ancestro de Ranunculales y todos sus parientes próximos ya extintos (el "stem group" Ranunculales) en unos 122-120 Ma, y la divergencia ya dentro del clado en unos 121-114 Ma. Todas las familias divergieron antes de los 105 Ma excepto la división entre Ranunculaceae y Berberidaceae que ocurrió hace unos 104-90 Ma (ver Anderson et al. 2005 para más detalles de diversificaciones posteriores dentro del clado).
Gleissberg y Kadereit (1999[29]) discuten la evolución de la morfología de las hojas en el clado, y consideran que las hojas de tipo ancestral (plesiomórficas) quizás hayan sido politernada, acrópeta, y basípetamente pedada. Los dientes de las hojas glandulares, con un claro, persistente, "swollen cap" (una especie de capuchón que tiene el diente, y que frecuentemente lo hace parecer un botoncito redondo, y suele esconder un hidatodo) al cual también llegan las venas laterales de órdenes más altos. Con respecto a los coléteres, Drinnan et al. (1994[30]) sugieren que la aparición de pétalos a partir de estambres debe haber ocurrido muchas veces dentro del clado, por ejemplo en Lardizabalaceae. Tamura (1965[31]) resume la evidencia por la naturaleza estaminodial de los pétalos en Ranunculaceae: los pétalos tienen un solo trazo foliar, están en los mismos parásticos que los miembros del androceo, y son similares a los estambres en su desarrollo temprano, y también usualmente son peltados (con pie). Con respecto a si la endexina del polen es lamelada, el APW remarca que las células de las antípodas normalmente no son simplemente persistentes; se puede encontrar una lista de datos en Williams y Friedman (2004[32]).
Ecología
Este clado, quizás especialmente Menispermaceae y Ranunculaceae, no es muy usado por las mariposas para alimentación de las polillas (Ehrlich y Raven 1964[33]). Hay datos acerca de una fertilización retrasada por dos meses o más después de ocurrida la polinización, en clados como Eupteleaceae, Circeasteraceae, Lardizabalaceae, y Ranunculaceae (ver Sogo y Tobe 2006[34] y referencias).
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Ranunculales.
Wikimedia Commons alberga una categoría multimedia sobre Ranunculales.
Referencias
- Ranunculales en: Stevens, P. F. 2001 Angiosperm Phylogeny Website, versión 7 (mayo 2006) y actualizado regularmente desde entonces. Fecha de acceso: 29 de abril del 2007.
- "Ranunculales". en: Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA. Capítulo 9, "Phylogenetic Relationships of Angiosperms".
Referencias citadas
- ↑ Angiosperm Phylogeny Group. 1998. An ordinal classification for the families of flowering plants. Ann Misouri Bot. Gard. 85: 531-553.
- ↑ The Angiosperm Phylogeny Group. 2003. "An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II". Botanical Journal of the Linnean Society, 141, 399-436. (pdf aquí )
- ↑ Stevens, P. F. 2001 Angiosperm Phylogeny Website, versión 7 (mayo 2006) y actualizado regularmente desde entonces.
- ↑ Chase, M. W. Soltis, D. E., Olmstead, R. G., Morgan, D., Les, D. H., Mishler, B. D., Duvall, M. R., Price, R. A., Hills, H. G., Qiu, Y.-L., Kron, K. A., Rettig, J. H., Conti, E., Palmer, J. D., Manhart, J. R., Sytsma, K. J., Michaels, H. J., Kress, W. J., Karol, K. G., Clark, W. D., Hedrén, M., Gaut, B. S., Jansen, R. K., Kim, K.-J., Wimpee, C. F., Smith, J. F., Furnier, G. R., Strauss, S. H., Xiang, Q.-Y., Plunkett, G. M., Soltis, P. S., Swensen, S. M., Williams, S. E., Gadek, P. A., Quinn, C. J., Eguiarte, L. E., Golenberg, E., Learn, G. H., Jr., Graham, S. W., Barrett, S. C. H., Dayanandan, S., y Albert, V. A. 1993. Phylogenetics of seed plants: An analysis of nucleotide sequences from the plastid gene rbcL. Ann. Missouri Bot. Gard. 80: 528-580.
- ↑ Drinnan, AN, PR Crane, y SB Hoot. 1994. Patterns of floral evolution in the early diversification of non-magnoliid dicotyledons (eudicots). Plant Syst. Evol. Suppl 8:93-122.
- ↑ Hoot, SB y PR Crane. 1995. Interfamilial relationships in the Ranunculidae based on molecular systematics. Plant Syst. Evol. Suppl. 9: 119-131.
- ↑ Loconte H, LM Campbell, y DW Stevenson. 1995. Ordinal and familial relationships of ranunculid genera. Plant Syst Evol. Suppl. 9: 99-118.
- ↑ Soltis, D. E. Kuzoff, R. K., Nickrent, D. L., Johnson, L. A., Hahn, W. J., Hoot, S. B., Sweere, J. A., Kuzoff, R. K., Kron, K. A., Chase, M. W., Swensen, S. M., Zimmer, E. A., Chaw, S.-M., Gillespie, L. J., Kresss, W. J., y Sytsma, K. J. 1997. Angiosperm phylogeny inferred from 18S ribosomal DNA sequences. Ann. Missouri Bot. Gard. 84: 1-49.
- ↑ Soltis DE, PS Soltis, ME Mort, MW Chase, V Savolainen, SB Hoot y CM Morton. 1998. Inferring complex phylogenies using parsimony: An empirical approach using three large DNA data sets for angiosperms. Syst Biol. 47: 32-42.
- ↑ Soltis DE, PS Soltis, MW Chase, ME Mort, DC Albach, M Zanis, V Savolainen, WH Hahn, SB Hoot, MF Fay, M Axtell, SM Swensen, LM Prince, WJ Kress, KC Nixon, y JS Farris. 2000. Angiosperm phylogeny inferred from 18S rDNA, rbcL, and atpB sequences. Bot. J. Linn. Soc. 133: 381-461. (resumen aquí)
- ↑ Källersjö M, JS Farris, MW Chase, B Bremer, MF Fay, ,CJ Humphries, G Petersen, O Seberg, y K Bremer. 1998. Simultaneous parsimony jacknife analysis of 2538 rbcL DNA sequences reveals support for major clades of green plants, land plants, and flowering plants. Pl. Syst. Evol. 213: 259-287.
- ↑ Hoot SB, S Magallón, y PR Crane. 1999. Phylogeny of basal eudicots based on three molecular data sets: atpB, rbcL, and 18S nuclear ribosomal DNA sequences. Ann. Missouri Bot. Gard. 86: 1-32.
- ↑ Savolainen V, MW Chase, SB Hoot, CM Morton, DE Soltis, CBayer, MF Fay, AY de Brujin, S Sullivan, y Y-l Qiu. 2000a. Phylogenetics of flowering plants based upon a combined analysis of plastid atpB and rbcL gene sequences. Syst Biol. 49: 306-362.
- ↑ Savolainen V, MF Fay, DC Alback, A Backlund, M van der Bank, KM Cameron, SA Johnson, MD Lledó, J-C Pintaud, M. Powell, MC Sheahan, DE Soltis, PS Soltis, P Weston, WM Whitten, KJ Wurdack, y MW Chase. 2000b. Phylogeny of the eudicots: a nearly complete familial analysis based on rbcL gene sequences. Kew Bull. 55: 257-309.
- ↑ a b c Cronquist, A. 1981. An integrated system of classificacion of flowering plants. Columbia University Press, Nueva York.
Error en la cita: Etiqueta
<ref>no válida; el nombre «Cronquist 1981» está definido varias veces con contenidos diferentes - ↑ a b Thorne RF. 1974. A phylogenetic classification of the Annoniflorae. Aliso 8: 147-209.
- ↑ a b Thorne RF. 1992. Classification and geography of the flowering plants. Bot Rev. 58: 225-348.
- ↑ Cronquist, A. 1968. The evolution and classification of flowering plants. Houghton Mifflin, Boston.
- ↑ a b Cronquist, A. 1988. The evolution and classification of flowering plants. 2ª edición. New York Botanical Garden, Bronx. Error en la cita: Etiqueta
<ref>no válida; el nombre «Cronquist 1988» está definido varias veces con contenidos diferentes - ↑ Dahlgren, R. M. T. 1983. General aspects of angiosperm evolution and macrosystematics. Nordic. J. Bot. 3: 119-149.
- ↑ Takhtajan, A. 1969. Floweringplants: Origin and dispersal. Smithsonian institution press. Washington, DC.
- ↑ Takhtajan, A. 1980. Outline of the classification of floweing plants (Magnoliophyta). Bot. Rev.'}' 46: 225-359.
- ↑ Donogue MJ y JA Doyle. 1989. Phylogeneticc analysis of angiosperms and the relationships of Hamamelidae. En: Evolution, systematics, and fossil history of the Hamamelidae, vol. 1, Introduction and "lower" Hamamelidae. Syst. Assoc. Special Vol. 40A, P. R. Crane y S. Blackmore (eds.), 17-45. Clarendon Press, Oxford.
- ↑ Doyle JA, MJ Donoghue y EA Zimmer. 1994. Integration of morphological and ribosomalRNA data on the origin of angiosperms. Ann. Missouri Bot. Garden81: 419-450.
- ↑ Árbol en APW
- ↑ Endress, PK. 1995. Floral structure and evolution in Ranunculanae. Plant Syst. Evol. Suppl. 9: 47-61.
- ↑ Magallón, S. A., Crane, P. R., y Herendeen, P. S. 1999. Phylogenetic pattern, diversity, and diversification of eudicots. Ann. Missouri Bot. Gard. 86: 297-372.
- ↑ Anderson, C. L., Bremer, K., y Friis, E. M. 2005. Dating phylogenetically basal eudicots using rbcL sequences and multiple fossil reference points. American J. Bot. 92: 1737-1748.
- ↑ Gleissberg, S., y Kadereit, J. W. 1999. Evolution of leaf morphogenesis: Evidence from developmental and phylogenetic data in Papaveraceae. Int. J. Plant Sci. 160: 787-794.
- ↑ Drinnan, Crane, y Hoot, S. B. 1994. Patterns of floral evolution in the early diversification of non-magnoliid dicotyledons (eudicots). Pp. 93-122, en: Endress, P. K., y Friis, E. M. (eds.), Early Evolution of Flowers. Springer, Vienna. [Plant Syst. Evol. Suppl. 8.]
- ↑ Tamura, M. 1965. Morphology, ecology and phylogeny of the Ranunculaceae IV. (Ranunculaceae of Eastern Asia: General Part IV). Sci. Rep. Osaka Univ. 14(1): 53-71.
- ↑ Williams, J. H., Jr., y Friedman, W. E. 2004. The four-celled female gametophyte of Illicium (Illiciaceae; Austrobaileyales): Implications for understanding the origin and early evolution of monocotyledons, eumagnoliids, and eudicots. American J. Bot. 91: 332-351.
- ↑ Ehrlich, P. R., y Raven, P. H. 1964. Butterflies and plants: A study in coevolution. Evolution 18: 586-608.
- ↑ Sogo, A., y Tobe, H. 2006. Delayed fertilization and pollen-tube growth in pistils of Fagus japonica (Fagaceae). American J. Bot. 93: 1748-1756.
{kind=link}