
Haptophyta
| Haptophyta | ||
|---|---|---|
 Pavlova sp. | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| (sin rango) |
Hacrobia | |
| Filo: | Haptista | |
| Subfilo: |
Haptophyta Hibberd 1972 ex Edvardsen & Eikrem in Edvardsen et al. 2000 | |
| Clases[1] y órdenes[2] | ||
| Sinonimia | ||
Coccolithophoridae, Lohmann 1902 | ||
Las haptofitas (Haptophyta o Prymnesiophyta) son un grupo de algas unicelulares de unas 500 especies, fundamentalmente marinas, que se caracterizan por la presencia de dos flagelos, un haptonema (apéndice contráctil superficialmente parecido a un flagelo) y células desnudas o cubiertas de placas calcáreas.[3][4][5] Incluye a los conocidos cocolitóforos, que tienen un exoesqueleto de placas calcáreas o cocolitos denominado cocósfera y constituyen el fitoplancton marino más abundante, especialmente en mar abierto, además de ser extremadamente abundantes como microfósiles. Algunas haptofitas periódicamente producen floraciones algales marinas tóxicas o generan una espuma desagradable que a menudo se acumula en las playas.
Ecología[editar]
Las haptofitas son un componente importante del fitoplancton marino y debido a su alta concentración juegan un papel importante como productores primarios en estos ecosistemas. En aguas tropicales son más abundantes, constituyendo alrededor del 45% del fitoplancton en las latitudes medias del Atlántico Sur. Disminuyen en abundancia hacia las latitudes templadas, aunque algunas haptofitas todavía se encuentran en aguas polares. Unas pocas especies viven en agua dulce o en el suelo. Muchas especies son simbiontes de otros organismos, por ejemplo, forman asociaciones con radiolarios.
Algunas especies periódicamente producen floraciones algales marinas tóxicas o generan una espuma desagradable que a menudo se acumula en las playas. Algunas especies segregan potentes exotoxinas que causan un gran mortalidad entre peces y moluscos.
Características[editar]


Las haptofitas son unicelulares, aunque a veces se agrupan en colonias o filamentos. El ciclo de vida generalmente comprende una sola etapa, pero en algunos casos alterna entre etapas flageladas y no flageladas. La etapa flagelada es unicelular y planctónica, mientras que la no flagelada suele ser colonial y bentónica. Las formas bentónicas pueden ser filamentosas, cocoides o palmeloides.[6]
Las haptofitas son organismos principalmente fotosintéticos, si bien algunos miembros son mixótrofos o heterótrofos. Muchas especies son fagotrofas y presentan vesículas en el citoplasma con las que digieren el material ingerido. No son selectivas en cuanto al material alimenticio que introducen en las vesículas y este puede consistir tanto de desechos orgánicos como de bacterias y otros pequeños organismos.[7]
Las células de las haptofitas tienen típicamente dos flagelos iguales o levemente desiguales, y lisos. La excepción es Pavlovales, donde uno de los flagelos es más largo que el otro y están cubiertos de pequeños pelos o escamas. Las haptofitas pueden nadar indistintamente con los flagelos dirigidos hacia delante o hacia la parte posterior del cuerpo. El movimiento es rápido y a menudo realizan cambios de dirección.
A menudo presentan un único orgánulo denominado haptonema, que es superficialmente similar a un flagelo pero que se diferencia del mismo en el conjunto de microtúbulos y en el uso. Su nombre proviene de las raíces griegas hapsis (tacto) y nema (hilo). Dependiendo de la especie, el haptonema puede ser largo o corto, y estar enrollado en espiral o no. Se encuentra situado entre los dos flagelos y no bate como estos, pero puede encogerse y estirarse y su función sería la fijación al sustrato, intervenir en el movimiento o apoyar la capacidad fagotrófica de algunas haptofitas. El haptonema está constituido por siete microtúbulos simples rodeados por la membrana del retículo endoplasmático.
Aunque se incluyen algunas formas con células desnudas (Pavlovales), la mayoría presentan células cubiertas de placas o escamas, de forma típicamente elíptica. Las placas orgánicas se originan en el aparato de Golgi y están a menudo envueltas en mucílago. Las placas pueden estar sin mineralizadar, consistiendo principalmente en carbohidratos, pero también pueden estar calcificadas, siendo muy raras las placas de sílice.
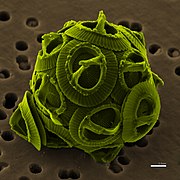
Las especies que tienen las placas calcificadas se conocen con el nombre de cocolitóforos. Estos tienen un exoesqueleto de placas calcáreas denominado cocolito y constituyen el fitoplancton marino más abundante, especialmente en mar abierto. La arquitectura de los cocolitos es muy variada, a menudo con una compleja ornamentación de espinas y rebordes, y se conocen especies con forma elíptica, pentágono, estrella, disco, etc. Los cocolitóforos se incluyen en los órdenes Coccolithales e Isochrysidales y son extremadamente abundantes como microfósiles. Estos se conocen desde el Carbonífero, hace unos 300 millones de años, y son muy abundantes durante el Jurásico.
Por lo general, las haptofitas presentan en cada célula de uno a cuatro cloroplastos discoidales alargados con un pirenoide en el centro del cloroplasto o como un abultamiento a un lado. Los tilacoides están agrupados de tres en tres y carecen de lamela periférica. Los cloroplastos están rodeados por cuatro membranas, estando la más externa cubierta de ribosomas y conectada con la membrana nuclear, formando el retículo endoplasmático cloroplástico. Como pigmentos fotosintéticos presentan clorofilas a, c1, c2 y c3 (esta última en Prymnesiophyceae). Como pigmentos accesorios presentan xantofilas de tipo dianinoxantina, diatoxantina y fucoxantina, además de β-caroteno. Como sustancias de reserva utilizan crisolaminarina, paramilo y gotas de lípidos, que se forman en vacuolas fuera del cloroplasto.
Se considera que los cloroplastos de las haptofitas proceden de la endosimbiosis secundaria de un alga roja. Los cloroplastos se pigmentan de forma similar a Ochrophyta (heterocontofitas, como en las algas doradas), pero se diferencian en el resto de la estructura celular, así que es posible que sean una línea evolutiva separada en la que los cloroplastos se deriven de endosimbiosis independientes.[8]
De la superficie de la célula se pueden originar seudópodos y filipodios, que pueden ser lineales o ramificados. Muchos son fagotrofos y en consecuencia tienen vacuolas alimenticias en el citoplasma con las que digieren bacterias y otros pequeños organismos. Presentan también un gran aparato de Golgi situado en el extremo anterior de la célula y, en algunos casos, también una vacuola contráctil. Algunos pocos géneros presentan una mancha ocular sensible a la luz, por ejemplo, Pavlova y Diacronema.[9] La mancha ocular se sitúa dentro o próximo al cloroplasto y consiste en gotitas de lípidos.[10] Las mitocondrias tienen crestas tubulares.
Las haptofitas usualmente se producen asexualmente por bipartición y la mitosis es abierta. La reproducción sexual puede ser común en algunos grupos.[2] Se ha encontrado algunas especies con un ciclo diplohaplonte, en la cual una etapa diploide flagelada alterna con una etapa haploide filamentosa. La etapa diploide es planctónica mientras que la haploide es bentónica.[6]
Especies notables[editar]
Ciertas especies como Emiliania huxleyi producen floraciones algales y algunos géneros como Chrysochromulina y Prymnesium producen toxinas que son fatales para los peces. Phaeocystis produce floraciones que generan una espuma desagradable que a menudo se acumula en las playas. Algunas haptofitas como Pavlova lutheri e Isochrysis son económicamente importantes, pues se utilizan extensivamente en la acuicultura.
Clasificación[editar]
Estudios moleculares y morfológicos dividen a Haptophyta en cinco órdenes agrupados en dos clases:[2]
- Pavlovophyceae. La superficie celular no presenta escamas, los flagelos son desiguales y a menudo están cubiertos de pelos. Algunas especies presentan una mancha ocular. Habitan las aguas costeras y salobres y a veces se encuentran en agua dulce. Solo se han descrito 13 especies en cuatro géneros, Pavlova, Diacronema, Exanthemachrysis y Rebecca y se consideran las haptofitas más primitivas.[10]
- Prymnesiophyceae. Superficie celular cubierta por escamas, los flagelos son iguales y lisos y la mancha ocular nunca está presente. Algunas especies presentan dos etapas en su ciclo de vida, alternando una etapa colonial o filamentosa con otra flagelada. Comprende la mayor parte de las especies de haptofitas, incluyendo los géneros bien conocidos Phaeocystis, Chrysochromulina, Prymnesium y los cocolitóforos, que periódicamente producen floraciones en costas y mar abierto, produciendo un impacto muy visible en el funcionamiento de los ecosistemas marinos.
Filogenia[editar]
Se han encontrado las siguientes relaciones:[11]
| Haptophyta |
| ||||||||||||||||||||||||
Origen[editar]
Las algas haptofitas se habrían originado por simbiogénesis entre una célula huésped hetrótrofa haptista y un alga endosimbionte heterokonta de acuerdo con la genética plastidial;[12] este endosimbionte sería una ocrofita de tipo Diatomista por la presencia de clorofila c3. Es por esto que haptofitas y ocrofitas tienen muchas similitudes, pues comparten la presencia de las clorofilas a, c1, c2, c3, β-caroteno y xantofilas como diatoxantina y fucoxantina, además de tener afinidades en la estructura de los tilacoides.[13]
Galería[editar]
-

-

-
-
 Floración de Emiliania huxleyi (Isochrysidales)
Floración de Emiliania huxleyi (Isochrysidales) -
 Floración de Phaeocystis (Phaeocystales)
Floración de Phaeocystis (Phaeocystales)





Referencias[editar]
- ↑ Caval.-Sm. et al. 2015
- ↑ a b c Adl, S.M. et al. (2012). The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514
- ↑ «www.ncbi.nlm.nih.gov».
- ↑ Satoh M, Iwamoto K, Suzuki I, Shiraiwa Y (2009). «Cold stress stimulates intracellular calcification by the coccolithophore, Emiliania huxleyi (Haptophyceae) under phosphate-deficient conditions». Mar. Biotechnol. 11 (3): 327-33. PMID 18830665. doi:10.1007/s10126-008-9147-0.
- ↑ «ITIS Standard Report». Consultado el 19 de julio de 2014.
- ↑ a b Hoek, C., Mann, D., & Jahns, H. M. (1995). Algae: an introduction to phycology. Cambridge university press.
- ↑ South, G. R., & Whittick, A. (2009). An Introduction to Phycology. John Wiley & Sons.
- ↑ R.A. Anderson (2004). Biology and Systematics of Heterokont and Haptophyte Algae. American Journal of Botany 91(10): 1508-1522.
- ↑ «Copia archivada». Archivado desde el original el 8 de diciembre de 2015. Consultado el 5 de agosto de 2015.
- ↑ a b Bendif, E. M., Probert, I., Hervé, A., Billard, C., Goux, D., Lelong, C., ... & Véron, B. (2011). Integrative taxonomy of the Pavlovophyceae (Haptophyta): a reassessment. Protist, 162(5), 738-761.
- ↑ El Mahdi et al. 2013, Erratum to: On the description of Tisochrysis lutea gen. nov. sp. nov. and Isochrysis nuda sp. nov. in the Isochrysidales, and the transfer of Dicrateria to the Prymnesiales (Haptophyta) Journal of Applied Phycology
- ↑ John W. Stiller et al. 2014, The evolution of photosynthesis in chromist algae through serial endosymbioses. Nature Communications 5, Article number: 5764 (2014) doi:10.1038/ncomms6764
- ↑ Robert A. Andersen 2003-2004 Biology and systematics of heterokont and haptophyte algae. Archivado el 11 de abril de 2016 en Wayback Machine. Am. J. Bot. October 2004 vol. 91 no. 10 1508-1522
| Control de autoridades |
|
|---|
 Datos: Q500531
Datos: Q500531 Multimedia: Haptophyta / Q500531
Multimedia: Haptophyta / Q500531 Especies: Haptophyta
Especies: Haptophyta
Enlaces externos[editar]
- Wikimedia Commons alberga una categoría multimedia sobre Haptophyta.